
Didelphis marsupialis Linnaeus, 1758
|
publication ID |
https://doi.org/ 10.1206/0003-0090(2001)263<0003:TMOPFG>2.0.CO;2 |
|
persistent identifier |
https://treatment.plazi.org/id/03B69D69-FFD8-3740-8687-FC31FF22FC9A |
|
treatment provided by |
Marcus |
|
scientific name |
Didelphis marsupialis Linnaeus |
| status |
|
Didelphis marsupialis Linnaeus View in CoL
VOUCHER MATERIAL: AMNH 266456– 266460 , 266462–266466 , 266468 , 266470 , 266471 , 266473 , 266475 , 267367 ; MNHN 1995.895 – 1995.901 . Total = 23 specimens (not including pouch young) .
IDENTIFICATION: All of our specimens of Didelphis from Paracou are referable to the large, blackeared species D. marsupialis , the type locality of which was restricted by Thomas (1911a) to Surinam. Our material agrees in all qualitative details with Husson’s (1978) description of Surinamese topotypes, but three of the four adults measured by Husson are larger than any collected at Paracou. To evaluate possible size differences, we borrowed Surinamese material for sidebyside comparisons with our vouchers; however, only four skinandskull preparations of adult specimens (all females) could be located. No differences in external characters were observed between Surinamese and French Guianan exemplars, and the measurement data we obtained (table 2) do not suggest any appreciable morphometric divergence. Considerable ontogenetic size variation is apparently characteristic of Didelphis species (Al len, 1901, 1902; Gardner, 1973), and it seems likely that Husson’s large specimens were just old animals that may have been preserved because of their unusual dimensions. The essential identity of the Surinamese and French Guianan material we examined supports the conclusions (hitherto undocument ed by published comparisons) of Thomas (1888) and Allen (1902) that D. karkinophaga Zimmermann and D. cancrivora Gmelin (both based on ‘‘Le Crabier’’, an opossum described by Buffon from Cayenne) are junior synonyms of D. marsupialis .
JulienLaferrière (1991) reported Didelphis albiventris and D. marsupialis as occurring syntopically in primary forest at Piste St.Élie (only 14 km WNW of Paracou), the first published record of the former species from French Guiana. Catzeflis et al. ( 1997) subsequently reported both species from primary forest near Petit Saut (about 28 km SSW of Paracou). In view of these records of sympatry from nearby localities, our failure to record the presence of D. albiventris at Paracou merits comment.
The diagnostic morphological characters of Didelphis marsupialis and D. albiventris are sufficiently striking that collected specimens cannot be misidentified by competent researchers alert to the possible presence of both species. In the Guiana subregion of Amazonia, these taxa are readily distinguished by facial markings (bolder in albiventris than in marsupialis ; Mondolfi and PérezHernández, 1984), ear color (the pinnae are usually tipped with white in albiventris but are entirely black in adult marsupialis ; op. cit.), caudal pelage (the furred basal portion of the tail is conspicuously longer in albiventris than in marsupialis ; M. D. Engstrom, personal commun.), and size ( albiventris is smaller; see measurements of the upper molar row of albiventris tabulated by Mondolfi and PérezHernández, 1984). Correlated molecular characters that may be useful for discriminating albiventris and marsupialis were recently discussed by Lavergne et al. ( 1997).
Although D. albiventris is definitely absent from our voucher material, we did not collect every individual Didelphis that we encountered at Paracou. Because many animals were sighted in dense vegetation where
TABLE 2 Measurements (mm) and Weights (g) of Adult Specimens of Didelphis marsupialis from French Guiana and Surinama
facial markings and other potentially informative details could not be distinguished, it is therefore possible that this species was seen but not recognized. Unfortunately, Didelphis virtually disappeared from our study area from 1992 to 1994, so we had little opportunity to collect specimens after we learned that D. albiventris could be expected to occur locally.
OTHER SPECIMENS EXAMINED: Surinam — Brokopondo, Brownsberg Nature Park ( CM 52697); Coronie, Totness ( CM 52702); Saramacca, Bigi Poika ( CM 52716, 52724). Listed specimens are adult skinandskull preparations; other Surinamese material examined ( CM, FMNH) are immature (subadults, juveniles) or incomplete (skull only/ skin only) specimens that are not useful for taxonomic comparisons.
FIELD OBSERVATIONS: All of our unambiguous records of Didelphis marsupialis from Paracou are based on collected specimens. Of these, 15 (65%) were trapped and 8 (35%) were shot. Nine specimens (39%) were trapped or shot in trees or other elevated substrates at heights ranging from 0.3 to 15 m above the ground, whereas the remainder were trapped or shot on the ground. Of 21 specimens accompanied by habitat data, 11 (52%) were taken in clearings, roadside sec ondary growth, or other disturbed habitats; the other 10 (48%) were taken in primary forest, at both welldrained and swampy sites. All specimens were shot or trapped at night, and all unvouchered sightings of Didelphis were also nocturnal. All encountered individuals were solitary; no collected female was carrying suckling young.
Gracilinanus emiliae (Thomas) Figures 11A, 11B View Fig , 12A View Fig , 17C, 17D View Fig , 18C, 18D View Fig
A single specimen from Paracou (AMNH 267006) is the first of this widespread but rarely collected species to be reported from French Guiana. Because the brief accounts by Thomas (1909), Tate (1933), Husson (1978), and Hershkovitz (1992) are inadequate for evaluating morphological similarities and differences with other congeners, we redescribe the species below.
TYPE MATERIAL: The holotype only, a male skin with skull and mandibles ( BMNH 9.3.9.10) collected by Emilie Snethlage on 13 February 1909 at ‘‘Para’’ (= Belém, formerly known as Para´), Brazil. The type was described by Thomas (1909: 379) as a ‘‘subadult’’, by Tate (1933: 189) as a ‘‘young adult’’, and by Hershkovitz (1992: 33) as a ‘‘juvenal’’. The animal is, in fact, very nearly adult with P3 and M4 both erupted but still a little below their adult positions in the toothrow.
Pine (1981: 59) suggested that the type locality should be construed as the state of Pará rather than the formerly eponymous city, but Thomas (always scrupulous about type localities) would surely have noted the lack of definite geographic information if he meant ‘‘Para’’ in the sense of a district larger than most European countries. A specimen collected at Capim (ca. 90 km ESE of Belém) provides independent evidence that the species may occur in the environs of the city.
DISTRIBUTION: Specimens that we examined document the presence of Gracilinanus emiliae in eastern Colombia, eastern Venezuela, southern Guyana, northern Surinam, northern French Guiana, and eastern Brazil (fig. 10). Several other published localities for this species are erroneous or unreliable (see Remarks, below).
EMENDED DESCRIPTION: Very small murine opossums (table 3) with smooth (not woolly) adult pelage; unruffled dorsal fur dull reddish brown, but basal twothirds dark gray; ventral fur pure white or cream from chin to groin (the hairs selfcolored, not grayish ba sally). Face marked by mask of black fur extending from mystacial pad to just behind outer canthus of eye on each side, and by narrow midrostral streak of pure orange fur; cheeks (below mask) white or creamcolored like throat; facial vibrissae (including long superciliary, mystacial, and genal hairs) mostly black (but some of the ventralmost genals are usually white); vibrissae of chin and throat (submental and interramal hairs) white. Ears not very large (just covering eye when laid forward over face), apparently naked (a sparse pelage of very short hairs is only visible under magnification), and very thin; opaque and pale basally (possibly yellow in life, but whitish in fresh alcoholic specimens); translucent and darker (brownish or grayish) distally. Gular glands (indicated by a naked or sparsely haired median patch of skin on the throat) present in all specimens examined, including one juvenile.
Wrists, ankles, and dorsal surface of feet covered with short pale (whitish or orangetinted) hairs. Manus and pes each with six plantar pads (thenar, hypothenar, and four interdigitals); thenar and first interdigital pads of manus separated by at least two rows of minute epidermal tubercles; thenar and first interdigital pads fused on pes of one fluid specimen ( AMNH 267006 ), touching but not fused on pes of two others ( ROM 35465, 35466 View Materials ) ; central palmar surface of manus smooth (not densely tubercular); claws of manual digits II–V small, not extending beyond fleshy apical pads. Scrotal epidermis of holotype entirely unpigmented, of another subadult ( AMNH 267006 ) with dark dorsal blotch surrounding suspensory stalk, of one fully adult specimen ( ROM 35466) entirely dark. At least nine (4–1–4) abdominalinguinal mammae present in one adult female ( ROM 35465) .
Tail much longer than headandbody (table 3); less than 1 cm furry at base; uniformly dark (grayish or brownish) without pale blotches, bands, or countershading. Caudal epidermis covered with very small scales in annular or spiral series, 3 numbering 40–50
3
The difference between spiral and annular arrangements of caudal scales cited by Tate (1933) as a useful character for diagnosing species groups of Marmosa (sensu lato) is less than obvious within Gracilinanus
TABLE 3 Measurements (mm) and Weights (g) of Subadult and Adult Specimens of Gracilinanus emiliae
rows/cm at middle of tail (counts from three fluid specimens and two dried skins). Median hair of triplet emerging from posterior margin of each scale about two scale rows long, thicker than lateral hairs, but not grossly flattened or petiolate.
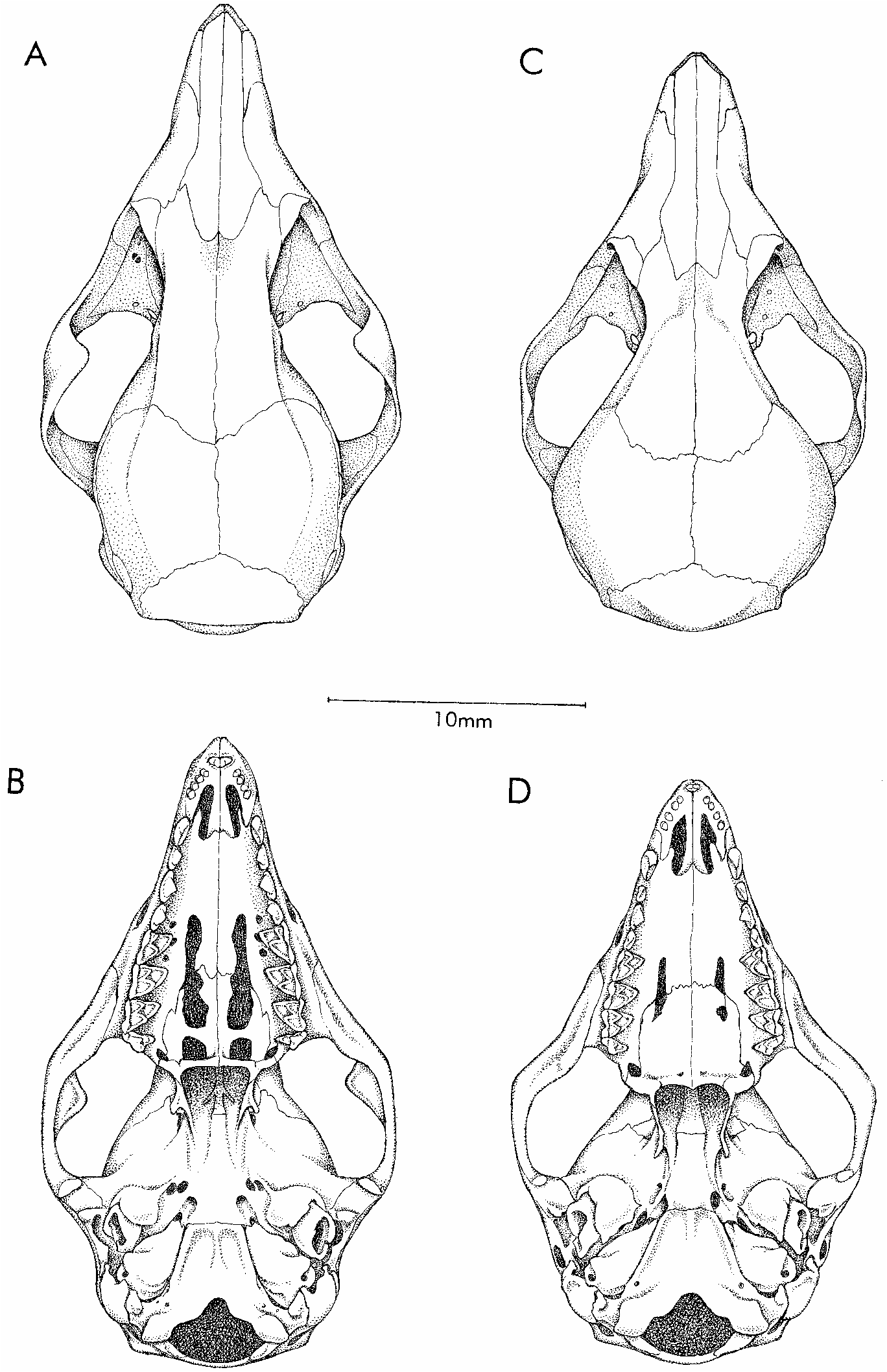
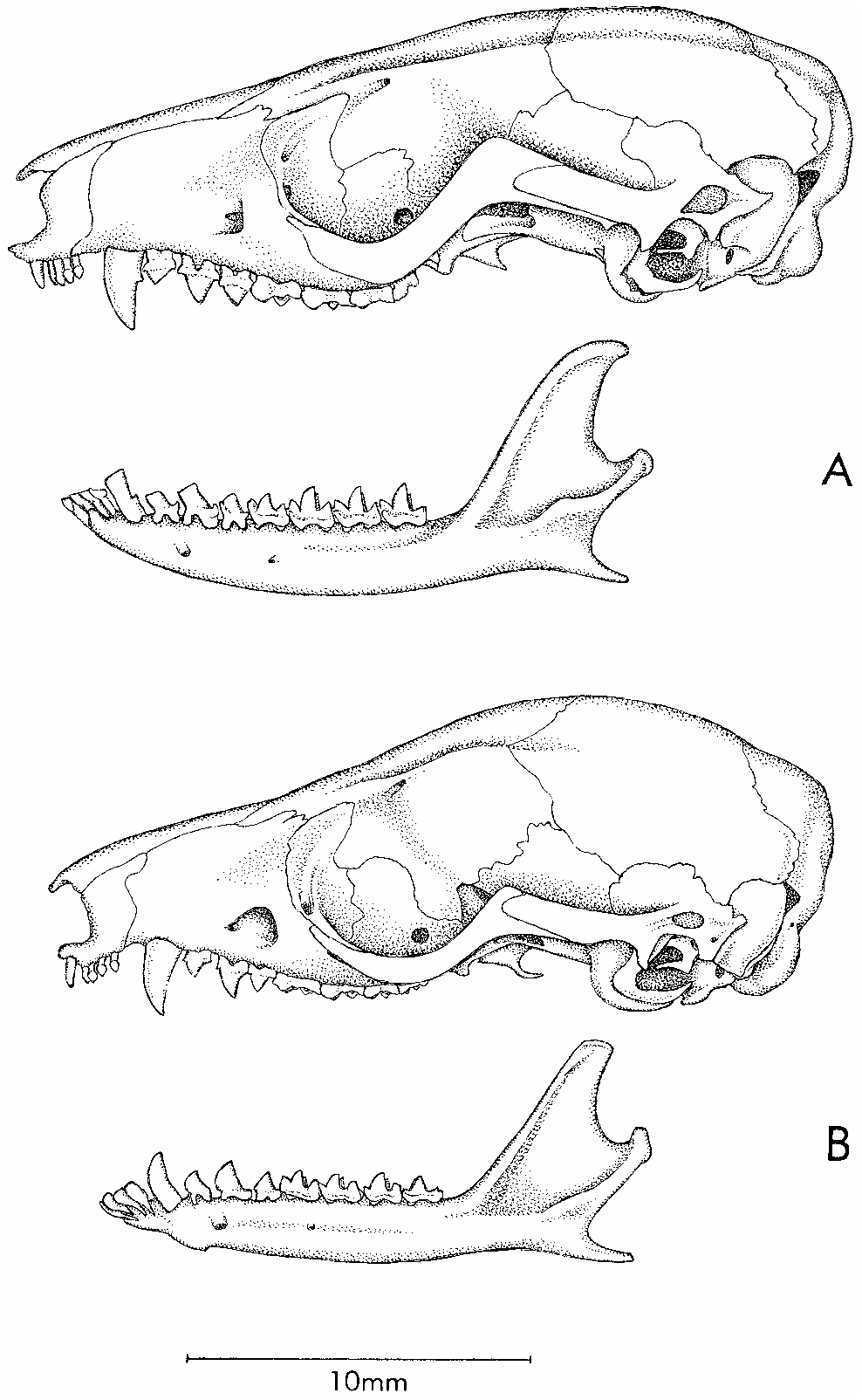
Skull (figs. 11, 12) with slender rostrum, incipiently beaded supraorbital margins, and absence of postorbital processes of frontals; orbits not conspicuously enlarged (delimited posteriorly by welldeveloped postorbital processes of jugals); braincase not greatly inflated, smooth and unmarked by prominent temporalis scars. Premaxillaries with small
(part of Tate’s microtarsus group). As noted by Hershkovitz (1992), spiral or annular series may occur on the same tail, and the perceived arrangement may be subject to observer bias. With the examples at hand of G. emiliae it is difficult to determine which, if either, descriptor is appropriate.
but distinct rostral process anterior to incisors; maxillarypremaxillary suture extending to posteriormost incisor alveolus; palate highly fenestrated, with large maxillopalatine and posteromedial (palatine) vacuities; one or more small maxillary palatal vacuities present unilaterally in one specimen ( AMNH 203363 ), bilaterally in three others ( BMNH 9.3 .9.10, ROM 35466, RMNH 18231 View Materials ). Alisphenoid wing of auditory bulla with welldeveloped anteromedial process bridging foramen ovale .
Upper incisors 2–5 subequal (not increasing in size from front to back); upper canine with small posterior accessory cusp; lower canine procumbent and premolariform (with flattened bladelike apex and small posterior accessory cusp); deciduous third premolars (dP3/dp3) large and molariform (figs. 17, 18); permanent third upper and lower premolars (P3/p3) slightly smaller than second premolars (P2/p2); metacones much larger than paracones on M1–3.
COMPARISONS: Gracilinanus emiliae and a sympatric taxon originally described by Hershkovitz (1992) as G. kalinowskii are similar in size and coloration (both have reddishbrown dorsal fur and pure white or creamcolored ventral fur), but they differ in many other characters as explained in the following account. Marmosa lepida (not known to occur at Paracou but reported from other localities in French Guiana and Surinam; appendix 1) is also externally similar to G. emiliae . Although adults of M. lepida are much larger than G. emiliae , immature specimens of the former species might be almost impossible to distinguish from the latter in the field. Cranially, adult M. lepida (see Husson, 1978: pl. 11) can be identified by their (1) distinctively longer premaxillary rostral processes, (2) prominent postorbital frontal processes, (3) lack of posteromedial palatal vacuities, (4) lack of anteromedial bullar processes, and (5) lack of mastoid exposure between the squamosal and parietal bones. Skulls of juvenile M. lepida that lack postorbital frontal processes can still be distinguished from G. emiliae by rostral, palatal, and bullar morphology, and by tooth size (see under Remarks, below).
REMARKS: Originally described as Marmosa emiliae (Thomas, 1909) , this species was subsequently referred to Didelphis (subgenus Grymaeomys ) by Matschie (1916), and to Marmosa (subgenus Thylamys ) by Cabrera (1958). The current allocation of emiliae to Gracilinanus follows Gardner and Creighton (1989).
Gracilinanus longicaudus , named by Hershkovitz (1992) from a single specimen (FMNH 87924) collected in eastern Colombia, does not differ significantly from G. emiliae in any external, cranial, or dental character. According to the original description, ‘‘... the combination of small size, long tail, whitish underparts, incomplete eye ring, and narrow skull separates [ longicaudus ] from all other described species [of Gracilinanus ]’’ (op. cit.: 39). However, the black mask of FMNH 87924 does, in fact, completely encircle the eye (a narrow border of black hairs is continuous below the lower lid and around the posterior canthus), and the other characters cited as diagnostic for longicaudus are matched by the type (and other referred specimens) of emiliae . Our sidebyside compar isons of AMNH 203363 (an adult male skin and skull from Capim, Brazil) with the holotype of emiliae in London and, later, with the holotype of longicaudus in Chicago, revealed no differences beyond those that might be expected among individuals from a single local population (for measurements, see table 3). We therefore regard G. longicaudus as a junior synonym of G. emiliae .
Eisenberg (1989) suggested that Gracilinanus emiliae might be conspecific with G. microtarsus from the Atlantic coastal rainforest of southeastern Brazil, but it would be difficult to select two more dissimilar congeners for comparison. Among other differences, Gracilinanus microtarsus is much larger and has a proportionately much shorter tail (see measurements in Tate, 1933; Hershkovitz, 1992), the ventral fur is buffy and graybased, the supraorbital margins are not beaded, and the crowns of I2–5 increase in size from front to back.
Gardner and Creighton (1989), Hershkovitz (1992), and Gardner ( 1993) listed Marmosa agricolai Moojen (1943) as a synonym of Gracilinanus emiliae , but Moojen’s illustration (op. cit.: fig. 1) and description are difficult to reconcile with this decision: the ratio of tail to headandbody for agricolai is only 1.28, the maxillarypremaxillary suture (as drawn) does not extend anteriorly to I5, the frontals are shown without supraorbital beads, the zygomatic arches are widely flared, the illustrated bullae appear to lack anteromedial processes, the upper canine appears to have a small anterior accessory cusp, and the third upper premolar (as drawn) is larger than the second. The type of agricolai (in Rio de Janeiro) should be reexamined to evaluate the true status and relationships of this nominal species.
We examined the two Brazilian specimens identified by Patterson (1992) as Gracilinanus emiliae and found them to be immature examples of Marmosa lepida . Both skulls have very long rostral processes of the premaxillae, lack posteromedial palatal vacuities, and their rounded alisphenoid bullae lack anteromedial processes; the erupted elements of the molar dentition are also larger than those of any specimens of G. emiliae but match the homologous teeth of adult M. lepida from eastern Peru (e.g., AMNH 78001, 98656). Because immature Marmosa lepida can be confused with Gracilinanus emiliae in size and external appearance, literature records of the latter (e.g., ÁvilaPires, 1964) should be regarded as suspect until diagnostic characters can be reconfirmed from specimens.
OTHER SPECIMENS EXAMINED: Brazil — Para´, Belém (BMNH 9.3.9.10, type), Capim (AMNH 203363). Colombia — Meta, Los Micos (FMNH 87924, type of Gracilinanus longicaudus ). Guyana — Upper Takutu Upper Essequibo, 12 km E Dadanawa (ROM 35465, 35466), no other locality data (ROM 33807). Surinam — Marowijne, Langamankondre (RMNH 18231). Venezuela — Monagas, 47 km SE Maturín (USNM 385066).
FIELD OBSERVATIONS: Our single specimen from Paracou was shot at 22:18 hours on 21 October 1992 as it perched about 4 m above the ground in dense secondary growth along a dirt road through welldrained forest (fig. 13).


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |