
Hyladelphys, VOSS & LUNDE & SIMMONS, 2001
|
publication ID |
https://doi.org/ 10.1206/0003-0090(2001)263<0003:TMOPFG>2.0.CO;2 |
|
persistent identifier |
https://treatment.plazi.org/id/03B69D69-FFD3-3775-86EA-FC04FD4CFE20 |
|
treatment provided by |
Marcus |
|
scientific name |
Hyladelphys |
| status |
gen. nov. |
Hyladelphys View in CoL , new genus
DIAGNOSIS: Very small didelphids distinguished from all other family members by the following combination of traits: four mammae in two abdominalinguinal pairs; dorsolateral margins of frontals beaded and strongly convergent anteriorly, without postorbital processes; premaxillae short, without rostral process anterior to incisor row; posteromedial (palatine) palatal vacuities absent; tympanic wing of alisphenoid without a welldeveloped anteromedial strut forming secondary foramen ovale; I2–5 not increasing in size from front to back, their crowns asymmmetrical and nonoverlapping; upper canine without anterior or posterior accessory cusps; P2 much larger than P3; deciduous premolars (dP3/dp3) very small and nonmolariform; molars not highly carnassialized (paracone smaller than metacone on M1–2, but paracone and metacone subequal on M3); lower canine not premolariform.
TYPE SPECIES: Gracilinanus kalinowskii Hershkovitz, 1992 .
CONTENT: Hyladelphys currently contains only the type species.
ETYMOLOGY: From ‘Ʋ´ λη (wood or forest), sometimes used in its Latinized form (hy laea) in reference to the predominant vegetation of the Amazonian lowlands (e.g., by Ducke and Black, 1953); and ƂeλϕƲ´ ς (womb), a traditional suffix for New World marsupial genera.
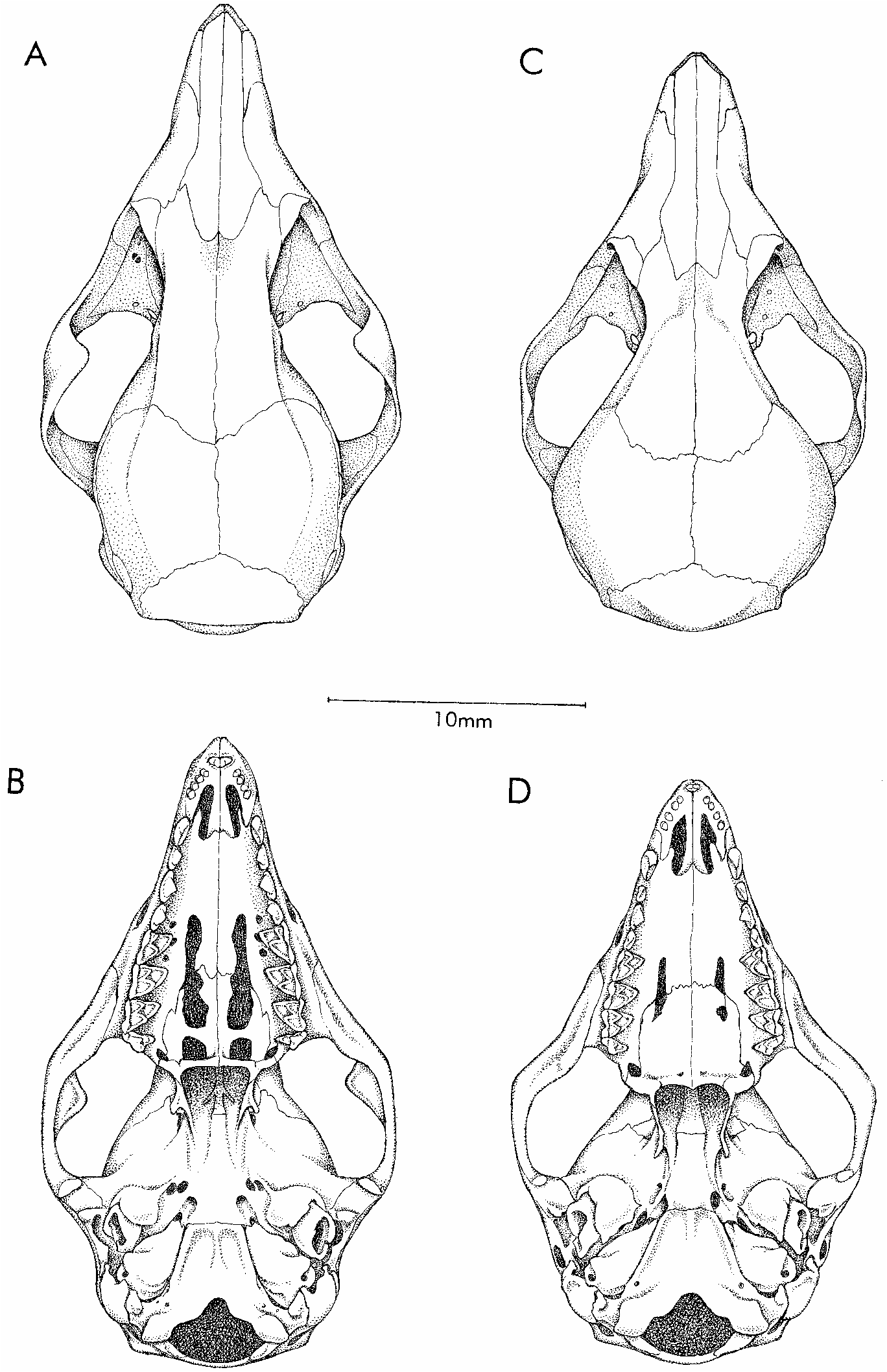
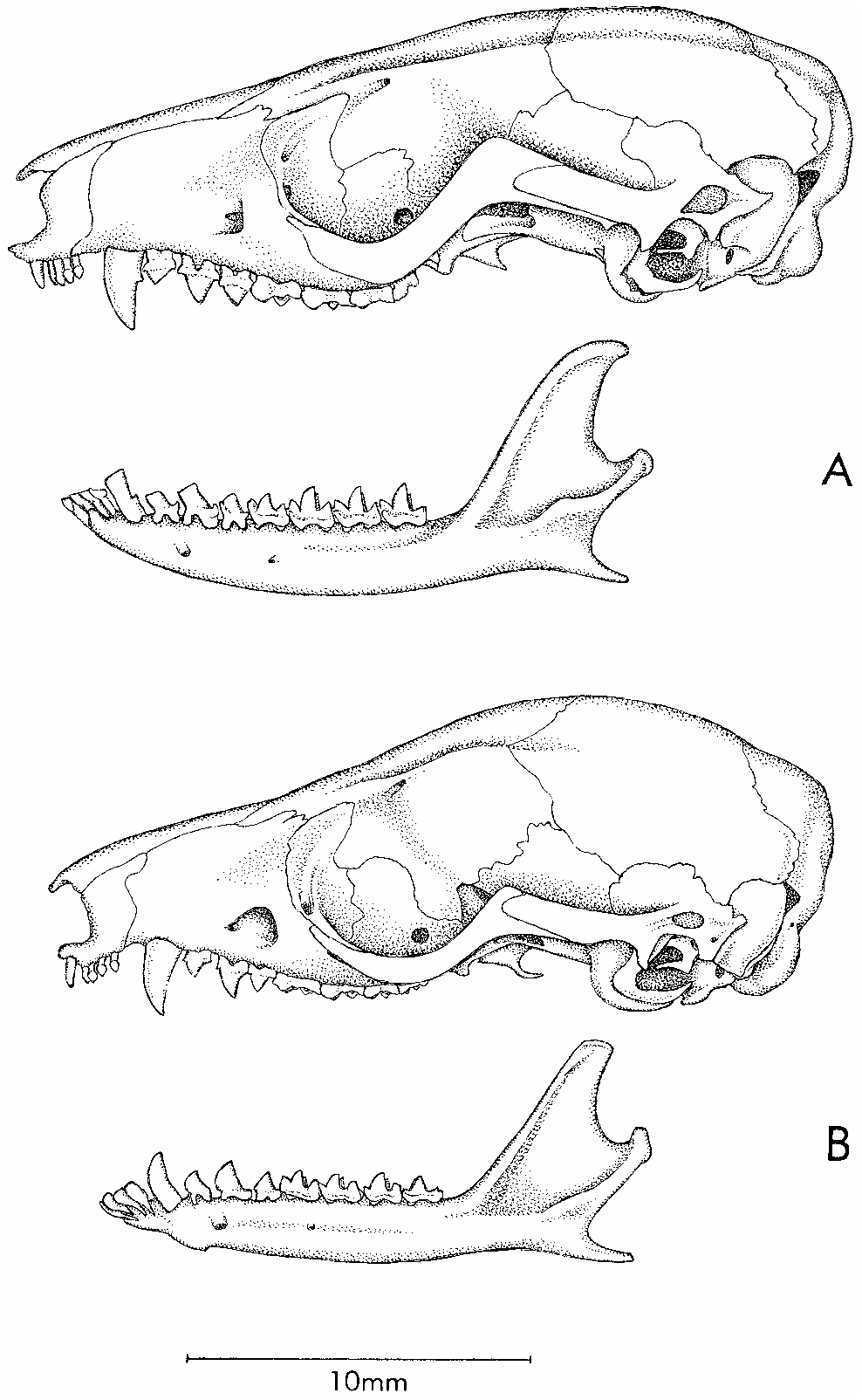
Hyladelphys kalinowskii (Hershkovitz) Figures 11C, 11D View Fig , 12B View Fig , 15A View Fig , 16A View Fig , 17A, 17B View Fig , 18A, 18B View Fig , 19B View Fig
Three specimens from Paracou ( AMNH 267338 , 267339 ; MNHN 1995.903 ) and one from Guyana (kindly loaned to us for identification by M. D. Engstrom) represent extraordinary range extensions of this distinctive species, originally described as Gracilinanus kalinowskii by Hershkovitz (1992) on the basis of two Peruvian specimens. We provide an emended description with comparisons and remarks based on this new material and our reexamination of Hershkovitz’s type series .
TYPE MATERIAL: The holotype, an adult female skin and skull ( FMNH 89991 View Materials ), collect ed on 9 July 1958 by Celestino Kalinowski at Hacienda Cadena (890 m elevation), Marcapata , Departamento Cuzco, Peru ; and the paratype, also an adult female skin and skull ( FMNH 65754 View Materials ), collected in October 1948 by J. M. Schunke at Chanchamayo (1100 m elevation), Departamento Junín, Peru .
DISTRIBUTION: Known only from Amazonian Peru, southern Guyana, and northern French Guiana (fig. 14).
EMENDED DESCRIPTION: Very small in all external and craniodental dimensions (table 4). Body pelage smooth (not woolly); dorsal fur unpatterned dull reddish brown, but basal twothirds of hairs dark gray; ventral fur pure white or cream from chin to groin (the hairs selfcolored, not grayish basally). Face boldly marked by broad mask of black fur extending from mystacial pad to base of ear on each side, and by prominent median streak of very pale orange fur extending from between eyes to rhinarium; cheeks (below mask) white like throat. Facial vibrissae (including long superciliary, mystacial, and genal hairs) mostly black (but a few of the ventralmost genals are white); vibrissae of chin and throat (submental and interramal hairs) white. Ears very large (covering eyes and extending to mystacial pads when laid forward over face), apparently naked (a sparse pelage of tiny hairs is visible only under magnification), and paperthin; opaque and bright yelloworange basally in fresh specimens (this color fading to white after a few months in alcohol), translucent and brownish distally. Eyes very large. Throat glands apparently absent. 4
4 Throat glands in murine opossums are indicated by a midventral gular patch of pigmented hair or bare skin (Tate, 1933). Although Hershkovitz (1992: 11) remarked that ‘‘neither marker is present in the two females of G. kalinowskii ’’, the paratype (FMNH 65754) was subsequently described as resembling the holotype externally ‘‘but with gular gland evident’’ (op. cit.: 38). We care
Wrists, ankles, and dorsal surface of metapodials covered with short, orange fur; manus and pes each with six plantar pads (thenar, hypothenar, and four interdigitals), the thenar and first interdigital touching but not fused on either manus or pes; central palmar surface smooth (not densely tubercular); claws of manual digits II–V large, extending beyond fleshy apical pads. Scrotum (of one adult male, AMNH 267338) pigmented,
fully examined the throat of FMNH 65754 under a dissecting microscope and found no external trace of glandular development. No discolored fur or bare patches are present on the throats of any of the four fluid specimens we examined from the Guianas.
TABLE 4 Measurements (mm) and Weights (g) of Adult Specimens of Hyladelphys kalinowskii
blue. Mammae 2–0–2 = 4, in two abdominalinguinal pairs (without an unpaired median teat; fig. 15). 5
Tail not very long and without furry base; dark (brownish) above and below, but indistinctly banded by absence of pigment over vertebral articulations. Caudal epidermis scaly beneath sparse covering of fine hairs; caudal scales in annular or spiral series, numbering about 25–35 rows/cm at middle of tail (counts from four fluid specimens). Median hair of triplet emerging from posterior margin of each scale usually about three scale rows long, thicker than lateral hairs but not grossly flattened or petiolate.
5 Hershkovitz (1992: 37) gave the mammary formula of kalinowskii as 7–1–7 = 15. This notation corresponds to what he described earlier (op. cit.: 11) as the ‘‘... prime formula for Gracilinanus ... of which the pectoral is 2–0–2 = 4, the abdominalinguinal, 5–1–5 = 11. The functional teat formula may be the same or less, depending on the number of attached young. Unused nipples are moreorless resorbed’’ (italics original). Contrary to this last assertion, however, unused teats remain everted and are not ‘‘resorbed’’ in adult female marsupials (Bresslau, 1920; TyndaleBiscoe and Renfree, 1987). Although unused teats are smaller than actively lactating teats, accurate mammary counts can be obtained even from anestrous adults. All of the five adult females of Hyladelphys kalinowskii that we examined have four welldeveloped mammae in two abdominalinguinal pairs; no other mammary loci were apparent, even with careful searching under high magnification.
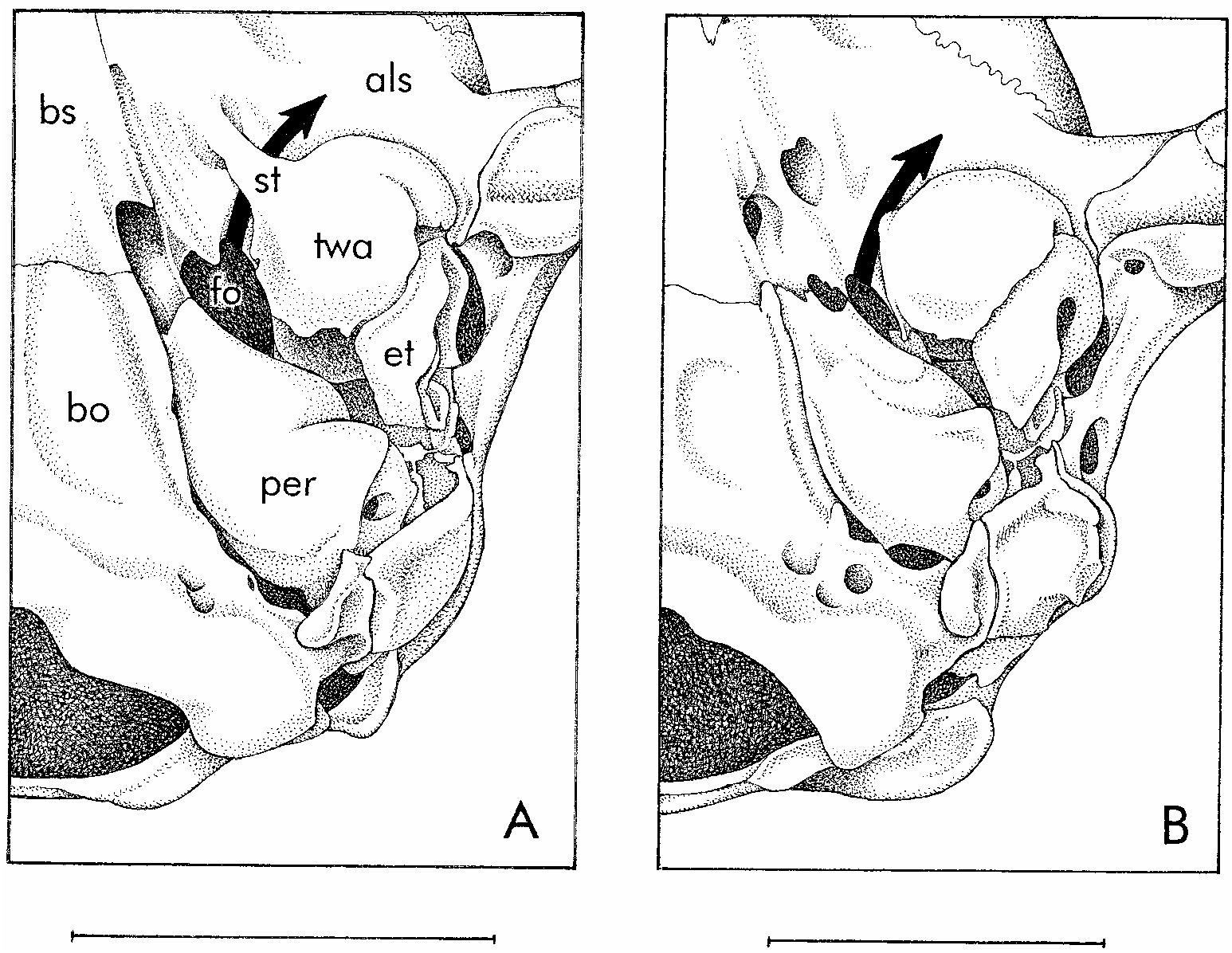
Skull (figs. 11, 12) with distinctively short, blunt rostrum; beaded and anteriorly convergent supraorbital margins; no postorbital frontal processes; very large orbits (their posterior limits indicated by weakly developed postorbital processes of the jugals); and laterally inflated braincase. Premaxillae without rostral process anterior to incisors; maxillarypremaxillary suture not extending to posteriormost incisor alveolus; maxillopalatine vacuities narrow, discontinuous in some specimens; posteromedial (palatine) and maxillary vacuities absent. 6 Alisphenoid wing of auditory bulla without a welldeveloped anteromedial process bridging the foramen ovale (see Remarks, below).
Upper incisors 2–5 subequal (not increasing in size from front to back), asymmetrical (the crowns with anterior but no posterior styles), and nonoverlapping (fig. 16); upper and lower canines without accessory cusps; lower canine erect and not premolariform; deciduous third premolars (dP3/dp3) extremely reduced, not molariform (figs. 17, 18); permanent upper and lower third pre
6 Hershkovitz (1992: 37–38) described the posteromedial vacuities of Hyladelphys kalinowskii as ‘‘small’’, but these fenestrae are absent in the holotype and the posterior palate of the paratype is destroyed. None of the specimens from Guyana and French Guiana have posteromedial vacuities.
molars (P3/p3) much smaller than second premolars (P2/p2); molars not highly carnassialized (metacone larger than paracone on M1 and M2, but metacone and paracone subequal on M3).
VARIATION: The four fluidpreserved spec imens (with extracted skulls) from Guyana and French Guiana agree in all qualitative characters with the Peruvian holotype despite several inconsistencies between Hershkovitz’s (1992) original description and that given above (see footnotes 4–6). The Peruvian specimens are slightly larger than French Guianan material (table 4), but the apparent size difference (perhaps best indexed by molar measurements) is no more than might be expected of conspecific samples collected almost 3000 km apart. The alisphenoid bullae of the holotype and of all four specimens from the Guianas lack anteromedial processes, but weakly developed processes (which are not fused to the floor of the braincase as they normally are in Gracilinanus , Marmosops , and Thylamys ) are present on the anteromedial aspect of both alisphenoid bullae of the paratype. We interpret this as intraspe cific variation because the paratype is not morphologically remarkable in any other respect.
COMPARISONS: Hyladelphys kalinowskii appears to differ from all other Recent didelphids in at least two qualitative characters. As far as is known, wellpreserved adult females of other didelphid taxa uniformly ex hibit oddnumbered mammary counts due to the presence of an unpaired median teat (fig. 15B); by contrast, caenolestids, microbiotheriids, and Old World marsupials have evennumbered mammary counts because their teats are bilaterally paired (Bresslau, 1920; Osgood, 1921, 1943; Tate, 1933, 1947, 1948). All reports of evennumbered didel phid mammary counts that we investigated were either field observations of lactating teats only (e.g., Hershkovitz, 1997: table 4), or were observations from specimens with midventral incisions that might have destroyed the median nipple (e.g., Marshall’s [1978a] count of four mammae from an FMNH skin of Glironia venusta ; W. T. Stanley, personal commun.). By contrast, our observations of identical (2–0–2 = 4) mam mary configurations in two Hyladelphys skins from Peru and two fluidpreserved specimens from French Guiana suggest that these are not rare variants or preservational artifacts. Instead, this taxon appears to be genuinely divergent from other didelphids in a character that displays considerable highertaxonomic stability among marsupials.
Hyladelphys also differs markedly from other didelphids in the morphology of its de
TABLE 5 Morphology and Size of Milk Premolars among Didelphids and Other Marsupials
ciduous dentition. Whereas previous descriptions of didelphid milk premolars have consistently reported these teeth as large and molariform (Flower, 1867; Thomas, 1888; Bensley, 1903; Tate, 1948; Archer, 1976b), there is in fact some substantial variation in the size and morphology of dP3/dp3 within the family (table 5). However, Hyladelphys is a conspicuous outlier: its milk teeth are much smaller than those of other confamilials, and they are uniquely nonmolariform in occlusal structure (figs. 17, 18). Instead, dP3/ dp 3 in Hyladelphys fall well within the range of milkpremolar morphologies seen in some Old World marsupial groups (Tate, 1947, 1948; Archer, 1976b; see illustrations in Luckett, 1993, 1994).
From superficially similar ‘‘marmosines’’, Hyladelphys differs in additional characters. Species of Gracilinanus , among which H. kalinowskii was previously classified, differ by having much smaller caudal scales (> 40 rows/cm), longer and narrower rostrums, less pronounced interorbital constrictions, smaller orbits, less inflated braincases, much more highly fenestrated palates, secondary foramina ovale formed by anteromedial struts of the alisphenoid tympanic wing, more carnassialized molars, and premolariform lower canines. Individual species of Gracilinanus differ from H. kalinowskii in other respects, but only G. emiliae merits explicit comparison here.
Gracilinanus emiliae occurs sympatrically with H. kalinowskii at Paracou, and because these species are similar in size and coloration (both have reddish dorsal fur and selfcolored whitish venters), they might be confused in the field. However, numerous external characters permit unambiguous discrimination. Gracilinanus emiliae differs from H. kalinowskii in facial markings (its black fa cial mask does not extend to the base of the ear, and the streak of orange fur between its eyes is darker and narrower) and has smaller ears, smaller manual claws (not extending beyond the fleshy digital pads), 4–1–4 = 9 mammae, and a much longer tail with small er scales and no trace of lighter banding over the vertebral articulations. Another external character that might distinguish G. emiliae and H. kalinowskii is the presence/absence of gular glands, but the variability of this trait within species is not well documented and, with so few examples available for comparison, its diagnostic value is uncertain. The skull of G. emiliae differs from that of H. kalinowskii in the cranial characters listed above for Gracilinanus and by the presence of a posterior accessory cusp on the upper canine (a trait not consistently exhibited by other congeners).
Members of other ‘‘marmosine’’ genera differ consistently from Hyladelphys by their larger size and sizecorrelated proportions (relatively longer rostrums, smaller orbits, less inflated braincases) and in the following qualitative contrasts (in addition to the mammary and milkdentitional traits mentioned previously): incrassate tails ( Thylamys ), grossly enlarged central hairs of each caudalscale triplet ( Marmosops ), distinct postorbital frontal processes ( Marmosa , Micoureus ), secondary foramina ovale ( Marmosops , Thylamys ), I2–5 conspicuously increasing in size from front to back ( Marmosops and some Marmosa ), P3/p3 larger than or subequal to P2/p2 ( Thylamys ), more carnassialized molars (all genera), and premolariform c1 ( Marmosops ). Insofar as can be inferred from polyprotodont character polarities (e.g., as hypothesized by Archer, 1976a, 1976b; Creighton, 1984; Reig et al., 1987; Wroe, 1997), Hyladelphys shows no clear pattern of synapomorphic resemblances with any other didelphid taxon. By the same token, evidence of a closer relationship to nondidelphid clades is also weak. In effect, our assignment of Hyladelphys to the family Didelphidae is based primarily on zoogeography and on morphological traits that are currently interpreted as marsupial plesiomorphies.
REMARKS: A published portrait of the head of Hyladelphys kalinowskii reconstructed from dried skins (Hershkovitz, 1992: fig. 14) is misleading in several details. The bulging eyes of our fluidpreserved specimens are proportionately about twice as large as those in the drawing. The pinnae in the illustration are shown bristling with short hairs, but the auricular pelage is actually microscopic and the unmagnified ears appear quite naked; the ears in life are also proportionately much larger than drawn. The portrait does not show the genal vibrissae, but these long black hairs are conspicuous against the short white fur of the cheeks in all of the specimens at hand. The mystacial vibrissae are depicted as fine, inconspicuous hairs that extend only to the outer canthus of the eye, but these robust whiskers actually extend nearly to the tips of the pinnae when laid back along the side of the head. The facial markings in the portrait also lack the vivid definition characteristic of this species: the mask is intensely black in fresh specimens and is boldly accentuated by a broad streak of very pale orange fur down the midline of the rostrum.
We examined the western Ecuadorean specimen (KU 135097) that Hershkovitz (1992: 42) identified as ‘‘ Gracilinanus sp. (new species)’’ and that he subsequently (op. cit.: 45) conjectured ‘‘... is most nearly like adult Gracilinanus kalinowskii ... ’’. The animal in question is a juvenile male (not a female as originally reported) preserved in fluid with an extracted skull, of which the first upper molar (crown length = 2.26 mm) suggests an adult size far larger than that of Hyladelphys or any known species of Gracilinanus . The large size of its caudal scales, absence of posteromedial palatal vacuities, and absence of an anteromedial alisphenoid strut bridging the foramen ovale are additional traits that cannot be reconciled with Gardner and Creighton’s (1989) diagnosis of Gracilinanus . From Hyladelphys, KU 135097 differs conspicuously by its welldeveloped rostral process of the premaxillae, welldeveloped postorbital jugal process, upper incisor morphology (I1–5 have large, overlapping, symmetrically rhomboidal crowns that increase in size from front to back), fully molariform dP3, and highly carnassialized molars. Based on these and other attributes, we refer this specimen to the genus Marmosa (sensu Gardner and Creighton, 1989) , within which it most closely resem
bles M. mimetra Thomas (1921) and other nominal taxa currently synonymized (Gardner, 1993) with M. robinsoni .
OTHER SPECIMENS EXAMINED: Guyana — East Berbice Corentyne, New River Falls ( ROM 34271) . Peru — Cuzco, Hacienda Cadena ( FMNH 8991 View Materials [type]) ; Junín, Chanchamayo ( FMNH 65754 View Materials [paratype]) ; Loreto, Río Gálvez, Nuevo San Juan ( MUSM 11031 ) .
FIELD OBSERVATIONS: Our first example of this species from Paracou (MNHN 1995.903) was shot as it perched on a palm frond about 1 m above the ground in swampy primary forest at 18:35 hours on 25 October 1992. The other two specimens (AMNH 267338, 267339) were taken from the same pitfall trapline, near a small stream in welldrained primary forest, on 21 August 1993.
Figures 7 View Fig , 19C View Fig , 21B View Fig
VOUCHER MATERIAL: AMNH 266416 , 266417 , 267368 , 267816 ; MNHN 1995.904 . Total = 5 specimens .
IDENTIFICATION: Our vouchers conform closely in all essential details of external and craniodental morphology with Tate’s (1933) and Husson’s (1978) authoritative descriptions of this species, the type locality of
which was restricted by Thomas (1911a) to Surinam. According to Tate (1933: 95), two subspecies are parapatrically distributed in Surinam and French Guiana: supposedly, Marmosa murina murina ranges ‘‘along the narrow coastal strip between the sea and the heavy rainforest’’, whereas ‘‘in the rainforest it is replaced by the smaller darker [M. m.] muscula [(Cabanis)].’’ Husson (1978), however, recognized only the former taxon in his Surinamese material, and all of the Surinamese specimens that we examined appear to represent a single recognizable form—closely conforming to Husson’s description and resembling our Paracou vouchers in qualitative characters and measurements (table 6)— that we assume to represent typical M. murina . Unfortunately, possible character differences between the types of murina and muscula are now difficult to evaluate.
Thomas (1892) identified two syntypes of Didelphys murina Linnaeus (1758) among the specimens of the Lidth de Jeude collection purchased by the BMNH in 1867. One of these (BMNH 67.4.12.542) was designat ed as the lectotype by Husson (1978), but his choice was unfortunate: the lectotype is a fluidpreserved adult female from which the skull has been extracted and apparently lost. The skull of the adult male paralectotype
TABLE 6 Measurements (mm) and Weights (g) of Adult Marmosa murina from French Guiana and Surinam
(BMNH 67.4.12.541), however, has measurements that fall within the range of variation that we observed among recently collected specimens from Surinam (table 6). Tate (1933) described the pelage color of BMNH 67.4. 12.541 in considerable detail, but neither this specimen nor the lectotype are really suitable for subspecific color comparisons because their pigments may have faded after more than two centuries in alcohol.
The holotype of Marmosa murina muscula (type locality: ‘‘Caraiben Niederlassung Arrai am obern Pomeroon’’ [Cabanis, 1848: 778], PomeroonSupenaam, Guyana) is a juvenile specimen (with dP 3 in place and M4 unerupted) that consists of the skin and skull of an animal originally preserved in fluid. Because of its immaturity, and because the pelage is now faded from preservative and discolored by age, the type of muscula is likewise unsuitable for subspecific comparisons. A small series of skins from Kartabo, Guyana, that Tate (1933) identified as M. m. muscula, however, are dorsally somewhat darker than our Paracou vouchers, but have whiter venters with less extensive lateral zones of graybased fur. Whereas the dorsal color difference is consistent with Tate’s diagnosis of muscula versus murina , the ventral color difference is not. The only qualitative cranial character cited by Tate as diagnostic of muscula, the absence of dorsal grooves along the supraorbital ridges, is apparently useless for defining this taxon (as represented by Tate’s own identifications) inasmuch as all the fully adult skulls from the Kartabo series (AMNH 42908, 48135, 99983, 142807) have grooved supraorbital ridges. Given that (1) the other character differences between murina and muscula cited by Tate are indefinite, (2) that the relevant types have discolored pelage and are incommensurate in age, and (3) that the material we examined from Surinam and French Guiana shows no appreciable divergence in size or coat color between coastal and interior populations, it does not seem useful to recognize these subspecies as valid at the present time.
We also examined the types of other nominal taxa from the Guiana subregion of Amazonia currently treated as subjective synonyms of Marmosa murina , including klagesi Allen (1900), chloe Thomas (1907), roraimae Tate (1931), and duidae Tate (1931). All of these are chiefly distinguishable by pelage characters—subtle differences in fur color, length, and texture—that perhaps vary clinally with environmental conditions as suggested by Tate himself (1933, 1939), who ranked them as no more than subspecifically distinct. By contrast, the conspecificity of some taxa from outside the Guiana subregion that are currently referred to M. murina (e.g., by Gardner, 1993) is more problematic. For example, M. quichua Thomas (1899a) from western Amazonia is craniodentally distinctive (Tate, 1933; personal obs.) and no justification for its synonymy with M. murina has apparently been published. In view of the lack of any critical review of the specieslevel taxonomy of Marmosa (sensu stricto) since Tate’s 1933 monograph, that work should still be considered the primary authority for species limits until compelling evidence is provided for alternative classifications.
OTHER SPECIMENS EXAMINED: French Guiana —Arataye (MNHN 1981.172, 1981.173, 1982.597, 1986.125), Cayenne (MNHN 1986.1024, 1986.1025), Les Nouragues (MNHN 1998.307), Piste St.Élie (MNHN 1981.417–1981.419, 1981.421, 1981.422, 1982.598), Saül (MNHN 1982.596, 1986.484). Guyana —‘‘Demerara River 29 miles above Georgetown’’ (BMNH 7.6.20.16 [holotype of chloe]); Cuyuni Mazaruni, Kartabo (AMNH 42907, 42908, 48135, 99983, 142807); Pomeroon Supenaam, ‘‘Caraiben Niederlassung Arrai am obern Pomeroon’’ (ZMB 2331 [holotype of muscula]). Surinam — Brokopondo, Finisanti (FMNH 95315–95319, 95321–95326, 95328); Marowijne, Oelemarie (CM 76729); Para, Zanderij (CM 68346, 68353); Saramacca, Raleigh Falls (CM 68354, 68355, 68356); Suriname, Lelydorpplan (FMNH 95329, 95330, 95332). Venezuela — Amazonas, Mt. Duida (AMNH 76984 [holotype of duidae]); Bolívar, Ciudad Bolívar (AMNH 16121 [holotype of klagesi]), Arabupu (AMNH 75703 [holotype of roraimae]). Without locality data —(BMNH 67.4.12.541 [paralectotype of murina ], 67.4.12.542 [lectotype of murina ]).
FIELD OBSERVATIONS: All of our records of Marmosa murina at Paracou are based on specimens collected in secondary growth and other manmade habitats: two were shot at night as they perched 1–2 m above the ground in dense roadside vegetation, one was caught by hand at night in a garden, and two were found killed and partially eaten (probably by domestic cats) near houses in clearings.






| ROM |
Royal Ontario Museum |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |