
Peltodoris, BERGH, 1880
|
publication ID |
https://doi.org/ 10.1046/j.1096-3642.2002.00039.x |
|
persistent identifier |
https://treatment.plazi.org/id/03B5879A-7565-6C79-90BD-FDFC254BAE36 |
|
treatment provided by |
Carolina |
|
scientific name |
Peltodoris |
| status |
|
GENUS PELTODORIS BERGH, 1880 View in CoL
Peltodoris Bergh, 1880: 41 View in CoL . Type species: Peltodoris atromaculata Bergh, 1880 View in CoL , by subsequent designation by O’Donoghue (1929).
Phialodoris Bergh, 1889: 908 View in CoL . Type species: Phialodoris podotria Bergh, 1889 View in CoL , by monotypy, syn. nov.
Montereina MacFarland, 1905: 38 View in CoL . Type species: Montereina nobilis MacFarland, 1905 View in CoL , by original designation, syn. nov.
Diagnosis
Dorsum covered with simple tubercles, stiffened by integumentary spicules, which occasionally protrude from the dorsal surface in an irregular fashion. Head with two conical oral tentacles. Anterior border of the foot grooved and notched. Labial armature smooth. Radula composed of simple, hamate teeth. The outermost teeth may be simple or denticulate. Reproductive system with a flattened, granular prostate, having two well differentiated regions. Penis and vagina devoid of hooks. Vestibular or accessory glands absent.
Remarks
Bergh (1880) described the genus Peltodoris based on Peltodoris atromaculata Bergh, 1880 . Peltodoris is characterized by having the dorsum covered with tubercles, finger-like oral tentacles, tripinnate gill, labial armature without jaws, radula with simple, hamate teeth, large prostate and penis and vagina unarmed. Bergh (1880) distinguished Peltodoris from Discodoris on the basis of the harder body consistency and especially because of the lack of jaws.
Bergh (1889) introduced the new genus Phialodoris based on Phialodoris potrida Bergh, 1889 from Amboine. He regarded Phialodoris as very close to Discodoris and Peltodoris , and only distinguishable from the latter by the peculiar shape of the penis. The penis of Phialodoris potrida is cylindrical with a cup-shaped apex armed with very small cones. Other characteristics of this species are similar to those of other members of Peltodoris , including the absence of jaws. There is no doubt that Phialodoris is a synonym of Peltodoris , and the peculiar penis shape is probably due to a preservation artifact or interspecific variation.
MacFarland (1905) described the genus Montereina based on Montereina nobilis MacFarland, 1905 . The diagnostic features of this genus are firm body, tuberculate dorsum, long and conical tentacles, large gill, differentiated prostate and vagina and penis unarmed. No further species have been assigned to this genus, which was later synonymized with Anisodoris Bergh, 1898 by MacFarland (1906). According to Valdés & Gosliner (2001), the genus Anisodoris , which is a synonym of Diaulula Bergh, 1878 , is characterized by having the dorsum covered with caryophyllidia. The anatomy of M. nobilis is similar to that of species of Peltodoris , and both names are here regarded as synonyms. Other species from the Pacific coast of South America, such as Doris variolata d’Orbigny, 1837 , previously assigned to the genus Anisodoris , should also probably be transferred to Peltodoris .
Eliot (1906b) pointed out that Peltodoris only differs from Discodoris in lacking a labial armature (jaws) and it should be regarded as a subgenus of Discodoris . Later, Thompson (1975) synonymized Peltodoris with Discodoris with no justification. In the following years a few authors followed Thompson’s authority and cited the type species of Peltodoris in the binomen Discodoris atromaculata ( Cattaneo-Vietti et al., 1990) . However, most authors maintained the usage of Peltodoris as a valid genus ( Castiello et al., 1980; Barletta, 1981; Schmekel & Portmann, 1982; Jonas, 1986; Perrone, 1992; Ávila, 1996).
The phylogenetic analysis carried out in the present paper indicates that Discodoris and Peltodoris belong in two different clades (see below); therefore, the genus Peltodoris is retained as valid.
PELTODORIS ATROMACULATA BERGH, 1880 View in CoL
( FIGS 4H View Figure 4 , 28 View Figure 28 , 29 View Figure 29 )
Peltodoris atromaculata Bergh, 1880: 45–46 View in CoL .
Type material
SYNTYPE: Naples , Italy, spring of 1880, one specimen, 34 mm preserved length ( ZMUC GAS-2054 )
Additional material
Islas Medas, La Escala, west coast of Gerona , Spain, three specimens, 25–34 mm preserved length, leg. T . M. Gosliner ( CASIZ 099147 ). Cala Salada, Ibiza, Spain, one specimen, 49 mm preserved length, leg. A. Valdés ( CASIZ 119474 ). 1 km east of Caloura , Ilha São Miguel , Azores, Portugal, eight specimens, 44–67 mm preserved length, leg. T . M. Gosliner ( CASIZ 072584 ) .
External morphology
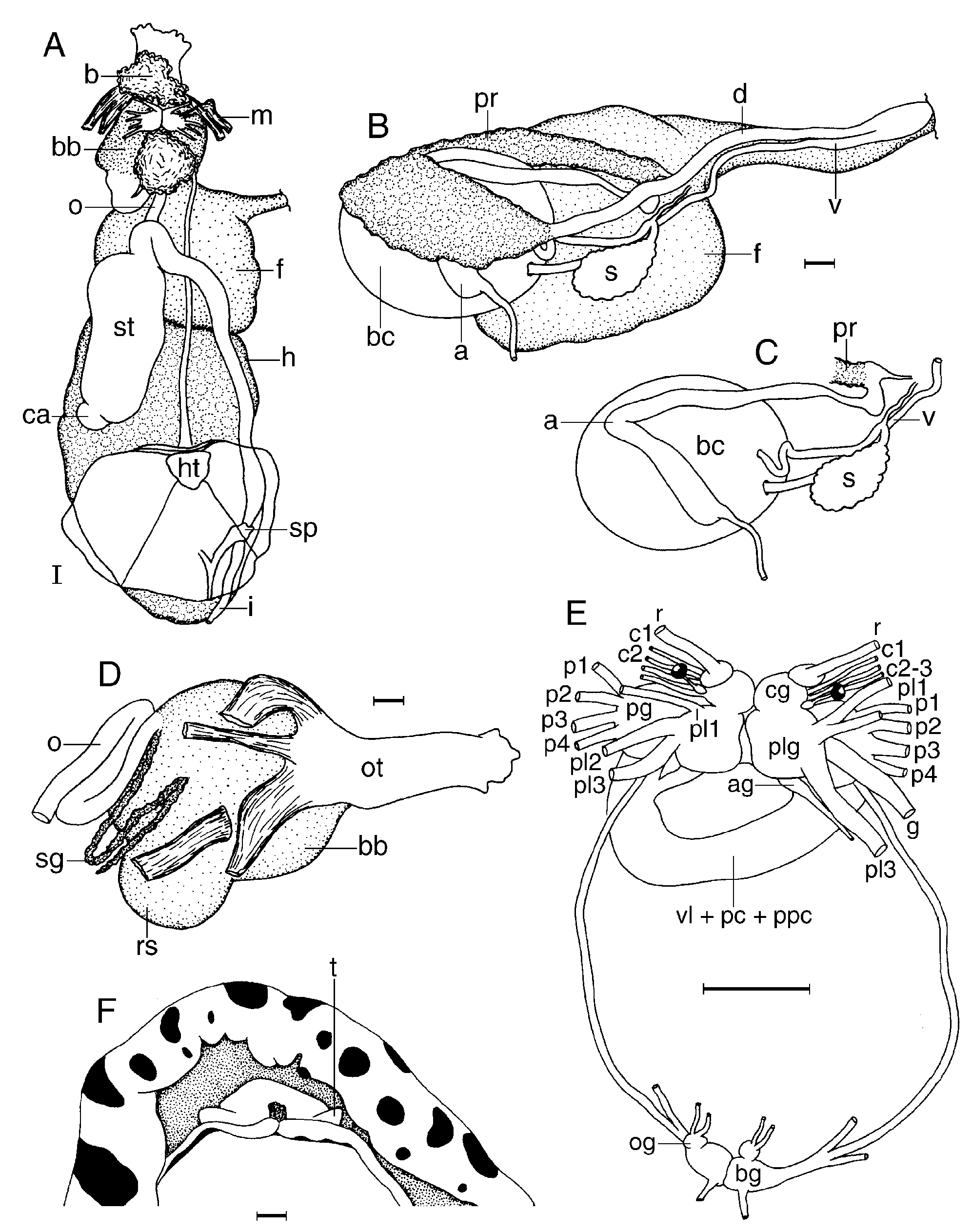
The general colour of the living animals is whitish to pale cream ( Fig. 4H View Figure 4 ). There is a number of dark brown or black large patches distributed on the dorsum, varying in shape and size. The rhinophores and gill are white or pale cream. The branchial leaves have some small dark brown or black spots. The whole dorsum is covered with small, conical tubercles, which have spicules protruding on their dorsal surface ( Fig. 28D View Figure 28 ). The largest tubercles are situated in the central region of the body. The rhinophoral and branchial sheaths have tubercles similar to those of the rest of the dorsum. There are six tripinnate branchial leaves, forming a circle. The anal papilla is situated in the centre of the branchial circle of leaves. The rhinophores are elongate, having 22 lamellae in a 52-mm preserved length specimen.
Ventrally there are two short and conical oral tentacles ( Fig. 29F View Figure 29 ). The anterior border of the foot is grooved and notched.
Anatomy
The posterior end of the glandular portion of the oral tube has six strong retractor muscles ( Fig. 29D View Figure 29 ) which attach to the body wall. The oval, muscular buccal bulb has two additional muscles attached; two long salivary glands connect with it at each side of the oesophageal junction. The buccal bulb is longer than the glandular portion of the oral tube. The labial cuticle is smooth. The radular formula is 22 ¥ 48.0. 48 in a 54-mm preserved length specimen. Rachidian teeth are absent. The inner lateral teeth are short, having a long, curved cusp and lacking denticles ( Fig. 28A View Figure 28 ). They also have a secondary, short and blunt cusp situated behind the main cusp. The teeth from the middle portion of the half-row are hamate, long and larger than those closer to the medial portion of the radula ( Fig. 28B View Figure 28 ). The outermost teeth are smaller and also smooth ( Fig. 28C View Figure 28 ). The oesophagus is short and connects directly to the stomach ( Fig. 29A View Figure 29 ).
The ampulla is long and thin, and branches into a short oviduct and the prostate ( Fig. 29C View Figure 29 ). The oviduct enters the female gland mass near to its centre. The prostate is flattened, long, folded and granular ( Fig. 29B View Figure 29 ), with two differentiated portions distinguishable by their colour and texture. It connects with a long duct that narrows and expands again into the small ejaculatory portion of the deferent duct. The muscular deferent duct opens into a common atrium with the vagina. The vagina is long. Near to its proximal end it joins the bursa copulatrix. From the bursa copulatrix leads another duct that connects to the seminal receptacle and the uterine duct. The bursa copulatrix is oval in shape, about 10 times as large as the seminal receptacle.
In the central nervous system ( Fig. 29E View Figure 29 ) the cerebral and pleural ganglia are fused and distinct from the pedal ganglia. There are four cerebral nerves leading from the left cerebral ganglion and three from the right one, and three pleural nerves leading from each pleural ganglion. There is a separate abdominal ganglion on the right side of the visceral loop. The buccal ganglia are near to the rest of the central nervous system, joined to the cerebral ganglia by two relatively long nerves. Gastro-oesophageal, rhinophoral and optical ganglia are present. The pedal ganglia are clearly separated, having four nerves leading from each one. The pedal and parapedal commissures are enveloped together with the visceral loop.
The circulatory system ( Fig. 29A View Figure 29 ) consists of a large heart and a two blood glands situated in front of and behind the central nervous system.
PELTODORIS NOBILIS ( MACFARLAND, 1905) View in CoL
( FIGS 4I View Figure 4 , 30 View Figure 30 , 31 View Figure 31 )
Montereina nobilis MacFarland, 1905: 38–39 View in CoL .
Type material
HOLOTYPE (by original designation): Monterey Bay, California, leg. F. M. MacFarland ( USNM 181284 About USNM ), not examined.
Additional material
Pacific Grove , Monterey Bay, California, USA, July– August 1923 and May 1926, 10 specimens, 24–67 mm preserved length, leg. F. M. MacFarland ( CASIZ 068237 ) .
External morphology
The general colour of the living animals varies from whitish to orange-yellow ( Fig. 4I View Figure 4 ). There is a number of dark brown or black small spots distributed on the entire dorsum below the level of the tubercles. The rhinophores have a light yellow base and a orange club. The gill is pale yellow with the apices of the leaves opaque white. The whole dorsum is covered with small, rounded tubercles ( Fig. 30D View Figure 30 ). The largest tubercles are situated in the central region of the body. The rhinophoral and branchial sheaths have tubercles no different from those on the rest of the dorsum. There are five tripinnate branchial leaves, forming a circle. The anal papilla is situated in the centre of the branchial circle of leaves. The rhinophores are elongate, having 17 lamellae in a 54-mm preserved length specimen.
Ventrally there are two long and conical oral tentacles ( Fig. 31E View Figure 31 ). The anterior border of the foot is grooved and notched.
Anatomy
The posterior end of the glandular portion of the oral tube has six strong retractor muscles ( Fig. 31D View Figure 31 ) which attach to the body wall. The oval, muscular buccal bulb has two additional muscles attached; two long and wide salivary glands connect with it at each side of the oesophageal junction. The buccal bulb is twice the length of the glandular portion of the oral tube. The labial cuticle is smooth. The radular formula is 27 ¥ 57.0. 57 in a 54-mm long specimen. Rachidian teeth are absent. The lateral teeth are narrow and elongate, having a single cusp and lacking denticles ( Fig. 30A View Figure 30 ). The teeth from the middle portion of the half-row are larger than those closer to the medial portion of the radula ( Fig. 30B View Figure 30 ). The outermost teeth are smaller and also lack denticles ( Fig. 30C View Figure 30 ). The oesophagus is short and connects directly to the stomach ( Fig. 31A View Figure 31 ).
The ampulla is very long and convoluted ( Fig. 31C View Figure 31 ). It branches into a short oviduct and the prostate. The oviduct enters the female gland mass near to its centre. The prostate is flattened and has two portions distinguishable by their colour and texture ( Fig. 31B View Figure 31 ). It connects with a long duct that narrows and expands again into the large ejaculatory portion of the deferent duct. The muscular deferent duct opens into a short common atrium with the vagina. The vagina is long and convoluted. At its proximal end it joins the bursa copulatrix. From the bursa copulatrix leads another duct connecting to the uterine duct and the seminal receptacle. The bursa copulatrix is oval in shape, about eight times as large as the seminal receptacle ( Fig. 31B View Figure 31 ).
In the central nervous system ( Fig. 31D View Figure 31 ) the cerebral and pleural ganglia are fused and distinct from the pedal ganglia. There are three cerebral nerves leading from each cerebral ganglion and two pleural nerves leading from each pleural ganglion. There is a separate abdominal ganglion on the right side of the visceral loop. The buccal ganglia are near to the rest of the central nervous system, joined to the cerebral ganglia by two relatively short nerves. Gastrooesophageal, rhinophoral and optical ganglia are present. The pedal ganglia are clearly separated, having four nerves each one. The pedal and parapedal commissures are enveloped together with the visceral loop.
The circulatory system ( Fig. 31A View Figure 31 ) consists of a large heart and a two blood glands situated in front of and behind the central nervous system.
Remarks
Peltodoris nobilis View in CoL is a well-known species from the Pacific Coast of North America (see McDonald, 1983). It was originally described in the genus Montereina ( MacFarland, 1905) View in CoL and later transferred to the genus Anisodoris View in CoL .




| ZMUC |
Zoological Museum, University of Copenhagen |
| T |
Tavera, Department of Geology and Geophysics |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
Peltodoris
| Valdés, Ángel 2002 |
Montereina
| MacFarland FM 1905: 38 |
Montereina nobilis
| MacFarland FM 1905: 39 |
Phialodoris
| Bergh R 1889: 908 |
Peltodoris
| Bergh R 1880: 41 |
Peltodoris atromaculata
| Bergh R 1880: 46 |