
Roboastra leonis, Pola, M., Cervera, J. L & Gosliner, T. M., 2005
|
publication ID |
https://doi.org/ 10.1111/j.1096-3642.2005.00167.x |
|
DOI |
https://doi.org/10.5281/zenodo.5113680 |
|
persistent identifier |
https://treatment.plazi.org/id/03B5622C-FF9D-1215-FEB0-84F1FE1EFD78 |
|
treatment provided by |
Carolina |
|
scientific name |
Roboastra leonis |
| status |
sp. nov. |
ROBOASTRA LEONIS View in CoL SP. NOV.
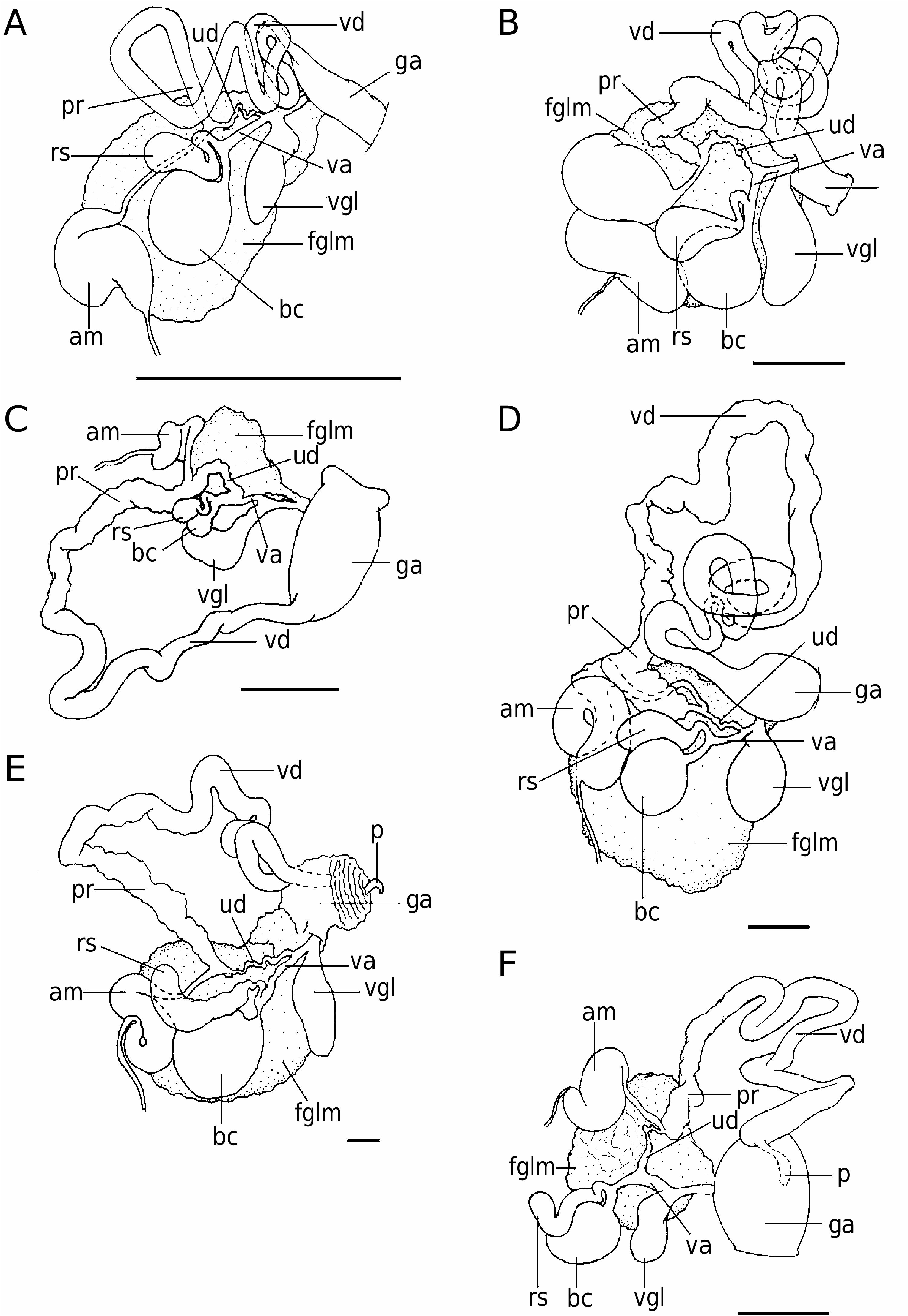
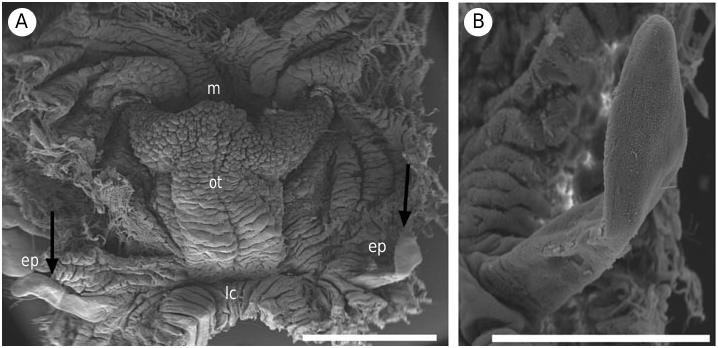
( FIGS 3F View Figure 3 , 4F View Figure 4 , 6E View Figure 6 , 7H View Figure 7 , 9 View Figure 9 , 10 View Figure 10 )
Roboastra View in CoL sp., Farmer, 1978: 384, fig. 20.
Roboastra View in CoL sp., Debelius, 1998: 181.
Type material: Holotype: Isla San Cristóbal, León Dormido, Kicker Rock , Galápagos Islands, Ecuador, May 1994, 1 specimen, 80 mm, collected by T. M. Gosliner aboard Mistral II, max. depth 26 m ( MECCD 4023 ) . Paratypes: same data as for holotype, 60 and 70 mm ( CASIZ 097548 ) . Other material: Isla Isabela, Punta Vicente Roca. Galápagos Islands , Ecuador, May 1994, 5 specimens, 25, 35, 45, 53 and 80 mm, collected by T. M. Gosliner aboard Mistral II, max. depth 23 and 31 m ( CASIZ 097577 ) . Isla Darwin, NE side, Galápagos Islands , Ecuador, May 1994, 1 specimen, 25 mm, collected by T. M. Gosliner aboard Mistral II, max. depth 18 m ( CASIZ 097554 ). The specimens were measured while preserved .
Etymology: Holotype found in the locality of León Dormido. It is also named leonis for its aggressive predatory habits.
Distribution: Currently reported from the Galápagos Islands, but Farmer (1978) included an illustration of a similar animal from La Paz (Gulf of California). Debelius (1998) reported it as endemic to the Galápagos Islands.
External morphology ( Fig. 6E View Figure 6 ): Body elongate and limaciform with a long and pointed posterior end of the foot. The preserved animals are 25–80 mm in length. The body surface is strongly wrinkled with numerous stripes of yellow ochre, green and bluishblack on the notum and both sides of the body. Some of the stripes are lighter in shade and can be continuous or discontinuous. The foot is linear and the edge of the foot is blue-black. The head is rounded with a pair of conical, completely retractile, perfoliate rhinophores with approximately 40 tightly packed lamellae. The base of the rhinophores is blue-black with a similar core; the leaves are bright orange and tipped in blueblack. The eyespots are brilliant blue. The oral tentacles are strongly developed and dorsolaterally grooved along part of their length and they are also blue-black. There are five nonretractile tripinnate gills; the three anteriormost gills are more highly developed. The gills form a semicircle surrounding the anal papilla, which is elevated and is blue-black in colour. The gills are blue-black distally and white proximally and the inner side of the rachis of each has a yellow line. The dark blue genital pore opens on the right side, midway between the gills and the rhinophores.
Internal morphology: Four specimens were dissected. The general arrangement of the internal organs is shown in Figure 9 View Figure 9 . The anterior digestive tract begins with a long, thick-walled muscular oral tube, which continues into the buccal mass. At their junction, a pair of slender elongate pouches opens into the digestive system ( Fig. 10A, B View Figure 10 ). The salivary glands are short and thick, opening on to the buccal mass and flanking the oesophagus. The labial cuticle is weak and lacks any armature. There is a well-developed blood gland, which is granular in texture. The radular formula of two 35-mm-long specimens (preserved) is 26 ¥ 4.1.1.1.4; that of the 60-mm-long specimen (preserved) is 32 ¥ 4.1.1.1.4 ( Fig. 7G View Figure 7 ). The rachidian tooth is broad, thin and very arched at its base, with two well-differentiated lateral cusps and a small one in the middle. The inner lateral tooth is very curved, having two very well developed cusps. The inner cusp is simple and larger than the outer one, which is very slender. The remaining lateral radular teeth are quadrangular, lack cusps or denticulation and become smaller near the margin.
The reproductive system is triaulic. That of one 45- mm-long specimen (preserved) ( CASIZ 097577) is shown in Figure 3F View Figure 3 . The genital mass is small compared with the total size of the specimens, but larger than in R. tigris . The hermaphroditic duct widens into a large S-shaped ampulla, which has thick walls. The ampulla narrows into the postampullary duct, which bifurcates into the vas deferens and oviduct. The short oviduct enters the female gland mass.
The deferent duct, which lacks a morphologically well-differentiated prostate, is long and coiled, ending in a dilated and darkly pigmented penial atrium. The duct has a uniform width, but is slightly narrower and thinner in the prostatic part. The penis is located within the distal end of this muscular portion and is armed with at least three different kinds of hooked and chitinous spines arranged in helicoidal rows. The types of spines and their arrangement on the penis are shown in Figure 4F View Figure 4 .
The bursa copulatrix and the seminal receptacle are well developed. The former is rounded and large, while the latter is elongate and smaller in size. The seminal receptacle has a short duct that connects to the vagina, near the bursa. The vagina is long and straight, opening into the genital atrium. The vaginal gland is very well developed, elongate, flattened, with muscular walls and joins the vagina at the distal part.
Remarks: Roboastra leonis is clearly distinguishable from other members of the genus. The external coloration is consistently different, while the number of lines on the notum and both sides of the body is higher than in the other species.
Roboastra luteolineata ( Baba, 1936) View in CoL also has many lines on its body, but the pigmentation of the gills, the rhinophores and most of the lines is different. There are also differences in the radular morphology of the two species. In R. leonis View in CoL the rachidian tooth is highly arched at its base, with two well-developed lateral cusps and a small one in the middle. In R. luteolineata View in CoL it is rectangular, with a slightly curved upper edge and three denticles (the central one is longer). Furthermore, the inner lateral tooth of R. leonis View in CoL has a simple inner cusp while in R. luteolineata View in CoL it is bifid.
R. tigris Farmer, 1978 View in CoL , which is biogeographically the closest species to R. leonis View in CoL , has a similar radular morphology, but its predominant body colour is greenish, yellow-ochre, or brown, with a few longitudinal dark navy blue stripes outlined in light green. These differences do not just involve coloration. In R. leonis View in CoL there are consistently more lines covering the body than in R. tigris View in CoL . Reproductive differences noted in the discussion of R. tigris View in CoL were consistent in the numerous mature specimens examined here.
Roboastra leonis View in CoL is the first species of the genus to have been found in the Galápagos Islands. It has been reported feeding on Tambja mullineri Farmer, 1978 View in CoL (Gosliner, unpublished data). Farmer (1978) provided a drawing of an animal similar to R. leonis View in CoL from La Paz, but the specimen was devoured by one of its R. tigris View in CoL companions in transit from the field to the laboratory. The external description of that species suggests that R. leonis View in CoL is also present in the Gulf of California; a photograph appears in Hickman & Finet (1999) and Debelius (1998).




| T |
Tavera, Department of Geology and Geophysics |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Roboastra leonis
| Pola, M., Cervera, J. L & Gosliner, T. M. 2005 |
Roboastra
| Debelius H 1998: 181 |
Roboastra
| Farmer WM 1978: 384 |