
Roboastra luteolineata, (BABA, 1936)
|
publication ID |
https://doi.org/ 10.1111/j.1096-3642.2005.00167.x |
|
DOI |
https://doi.org/10.5281/zenodo.5113672 |
|
persistent identifier |
https://treatment.plazi.org/id/03B5622C-FF94-120E-FF4E-83F9FE2FFDD4 |
|
treatment provided by |
Carolina |
|
scientific name |
Roboastra luteolineata |
| status |
|
ROBOASTRA LUTEOLINEATA ( BABA, 1936) View in CoL
( FIGS 3B View Figure 3 , 4B View Figure 4 , 6A View Figure 6 , 7A, B View Figure 7 , 8B View Figure 8 )
Nembrotha luteolineata Baba, 1936: 26–27 View in CoL , pl. 3, fig. 7; text-fig. 14.
Material examined: North coast, near Madang, Papua New Guinea, June 1992, 1 specimen, 24 m depth, 40 mm preserved, collected by T . M. Gosliner ( CASIZ: 086319). Ryukyu Island , Okinawa, Japan, March 1997, 1 specimen previously identified as Tambja affinis , 115 mm in life (70 mm preserved), 55 m depth, collected by R . F. Bolland from a bottom of mixed rock and coral rubble ( CASIZ: 115739). South Mulron Island , Western Australia, August 1995, 1 specimen previously identified as Tambja affinis , 45 mm preserved, collected by C.W. Bryce ( WAM: S23322 View Materials ) .
Distribution: Originally described from Japan ( Baba, 1936), this species has been also reported from New Zealand, Australia (Lord Howe Is., Heron Is.), Solomon Is., Kerama Is. (Okinawa, Japan), Thailand, Indonesia (Bali) and South Africa (Durban) ( Hamatani & Baba, 1976; Gosliner, 1987; Coleman, 1989; Debelius 1998; Ono, 1999, 2004; Suzuki, 2000; Armstrong & Armstrong, 2001; Coleman, 2001; Johnson, 2001; Takamasa, 2003; Nakano, 2004).
External morphology ( Fig. 6A View Figure 6 ): Body elongate and limaciform with a long and pointed posterior end of the foot. The preserved animals are 35–70 mm in length. The body surface is smooth or lightly wrinkled with numerous (6–8) yellow longitudinal stripes on either side of the body. A wide yellow submarginal band follows the inner notal edge. There are two stripes on the back, which pass posterior to the base of the gills. The stripes in the lateral parts of the back each arise from the outer corner of the rhinophores. In some specimens all these lines can also be green or orange or, more typically, a mix of bright yellow and green. There are two milky green markings between the rhinophores.
The ground colour is dark green or deep blue-black. The foot is linear and the edge of the foot is dark green or blue-black. The head is slightly expanded with a pair of conical, completely retractile perfoliate rhinophores with approximately 45 tightly packed lamellae. The oral tentacles are strongly developed and dorsolaterally grooved along a part of their length and they are also blue-black. There are five nonretractile tripinnate gills, with the three anteriormost being more highly developed. The gills form a semicircle surrounding the anal papilla, which is elevated and is also blue-black. The gills are dark, the same colour as the body, but the outer and inner sides of the rachis are green. The sheaths of the rhinophores, rhinophores, oral tentacles and pinnae of the gills are black. The genital pore opens on the right side, midway between the gills and the rhinophores.
Internal morphology: The anterior digestive tract begins with a long and muscular oral tube, that continues into the buccal mass. At the junction of the oral tube and the buccal mass a pair of large, thick, elongated pouches open into the digestive system. There are two small and elongated salivary glands on the buccal bulb, flanking the oesophagus. Green and small-developed blood gland, granular in texture, located above the intestinal loop. The buccal mass is shorter than the length of the oral tube. The labial cuticle is smooth. The radular formula of the 40-mmlong specimen (preserved) is 35 ¥ 3.1.1.1.3; that of the 115-mm-long specimen (in life) is 40 ¥ 4.1.1.1.4. The rachidian tooth is rectangular, with a slightly curved upper edge and three denticles; the central one is longer ( Fig. 7A View Figure 7 ). The inner lateral tooth typically has a strongly curved bifid inner cusp. The upper denticle of this inner cusp is slender and well developed, while the outer cusp is strong and well developed ( Fig. 7B View Figure 7 ). The outer lateral teeth are plate-like and become smaller near the margin ( Fig. 7A View Figure 7 ).
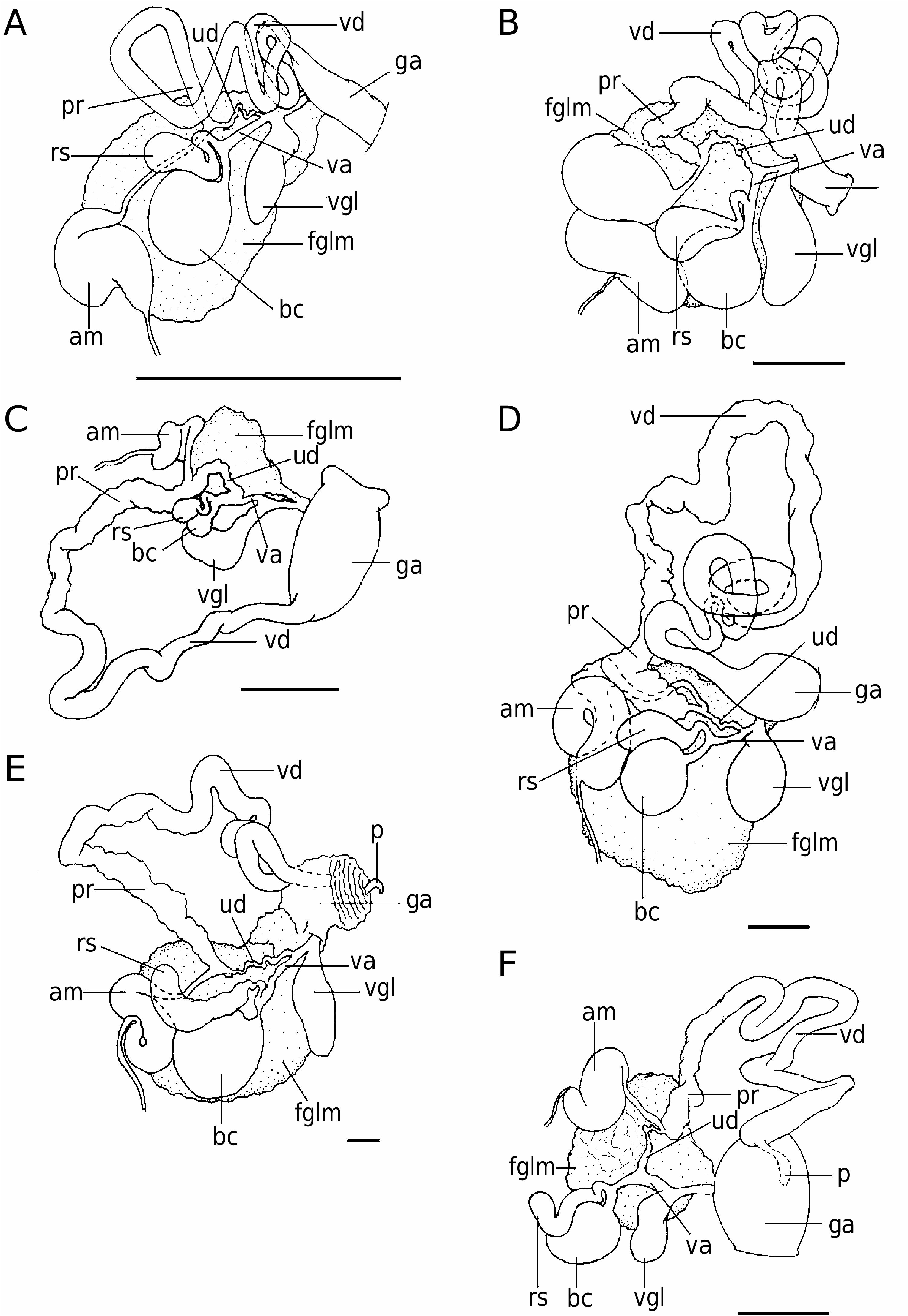
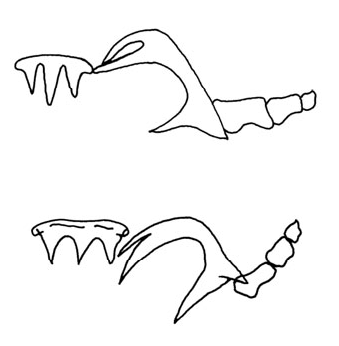
The hermaphroditic reproductive system is triaulic ( Fig. 3B View Figure 3 ). The genital mass fills the second quarter of the body cavity. The ampulla is large, folded thrice upon itself, with soft and thin walls. From its distal portion emerges the postampullary duct, which divides into the prostate and the oviduct. The prostate is wide, long and convoluted, distally narrowing and expanding again into the long muscular ejaculatory portion. The vas deferens is highly coiled. The distal portion of the deferent duct opens into a long, wide common atrium with the vaginal duct. The penis is located within the distal end of this muscular portion and it is armed with at least three different kinds of hooked and chitinous spines arranged in helicoidal rows ( Fig. 4B View Figure 4 ). The vaginal duct is thin, elongate and enters the large, spherical bursa copulatrix. Another duct connects the vagina to the smaller and pyriform seminal receptacle after making two loops. Distally to the bursa copulatrix, a short and coiled uterine duct leaves the vagina and enters the female gland mass. The vaginal gland is well developed, elongate and joins with the vagina before opening into the genital atrium.
Remarks: Roboastra luteolineata was described by Baba (1936) as Nembrotha luteolineata , based on a 30- mm-long specimen collected in Ishigakishima, Japan, and another specimen, 70 mm long, collected in the same locality. Baba (1936) gave two radular formulae (35 ¥ 3.1.1.1.3 and 40 ¥ 3.1.1.1.3) while Hamatani & Baba (1976) gave one: 36 ¥ 4.1.1.1.4. These results, together with the description of the external morphology, are almost entirely consistent with our description. The specimens studied here have five tripinnate gills instead of three ( Baba, 1936). Hamatani & Baba (1976) described five bipinnate gills, but we can see that they are tripinnate. The reproductive system of this species has not been previously described.
Roboastra luteolineata View in CoL has been identified in a number of popular books on Australian nudibranchs (Willan & Coleman, 1984; Coleman, 1989; Debelius, 1998) as Tambja affinis ( Eliot, 1904) View in CoL . However, that species, originally described from East Africa, was described as having a single median yellow stripe. Moreover, apart from other features, such as the milky green marking between the rhinophores, the radular teeth of R. luteolineata View in CoL and T. affinis View in CoL are very distinct. R. luteolineata View in CoL feeds on other polycerids, such as Tambja verconis (Basedow & Hedley, 1905) ( Armstrong & Armstrong, 2001) View in CoL , rather than the arborescent bryozoan Bugula dentata View in CoL , on which Tambja verconis View in CoL feeds ( Rudman, 1998b). Ono (1999, 2004) included photographs of R. luteolineata View in CoL feeding on another species of Tambja View in CoL / Nembrotha View in CoL in Japan.



| T |
Tavera, Department of Geology and Geophysics |
| R |
Departamento de Geologia, Universidad de Chile |
| WAM |
Western Australian Museum |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Roboastra luteolineata
| Pola, M., Cervera, J. L & Gosliner, T. M. 2005 |
Nembrotha luteolineata
| Baba K 1936: 27 |