
Dictyoplax undetermined
|
publication ID |
https://doi.org/10.1111/zoj.12138 |
|
persistent identifier |
https://treatment.plazi.org/id/03B48793-FFBD-F87C-CFEF-FEBD3F512C9B |
|
treatment provided by |
Carolina |
|
scientific name |
Dictyoplax undetermined |
| status |
|
DICTYOPLAX LECUS GEN. ET SP. NOV.
( FIGS 6 View Figure 6 , 7 View Figure 7 , TABLE 4)
Type material: Holotype, USNM 1110010 About USNM , MOV Johnson SeaLink I, dive 4905, 16 Nov. 2002, off Riding Rock , San Salvador, Bahamas, 24°03.5843′ N, 74°33.2534′ W, 762 m. GoogleMaps Paratypes, USNM 1231338 About USNM , MOV Johnson SeaLink I, dive 2306, 11 Dec. 1992, Plana Cay , Bahamas, 22°32.00′ N, 73°37.00′ W, 891 m GoogleMaps ; USNM 1231339 About USNM , MOV Johnson SeaLink I, dive 3408, 11 Dec. 1992, west of Riding Rock , San Salvador, Bahamas, 24°03.331′ N, 74°33.254′ W, 812 m GoogleMaps ; HBOM 002 View Materials :00028, MOV Johnson SeaLink I, dive 4507, 15 Nov. 2002, French Bay , San Salvador, Bahamas, 23°56.0048′ N, 74°30.9088′ W, 840 m GoogleMaps ; HBOM 002 View Materials :00029, MOV Johnson SeaLink I, dive 4622, 16 Oct. 2003, Plana Cay , Bahamas, 22°33.3592′ N, 73°37.2976′ W, 785 m GoogleMaps ; USNM 1231340 About USNM , MOV Johnson SeaLink I, dive 4623, 16 Oct. 2003, Samana Cay , Bahamas, 23°02.7928′ N, 73°45.6564′ W, 880 m. GoogleMaps
Diagnosis: As for the genus, as it is monospecific.
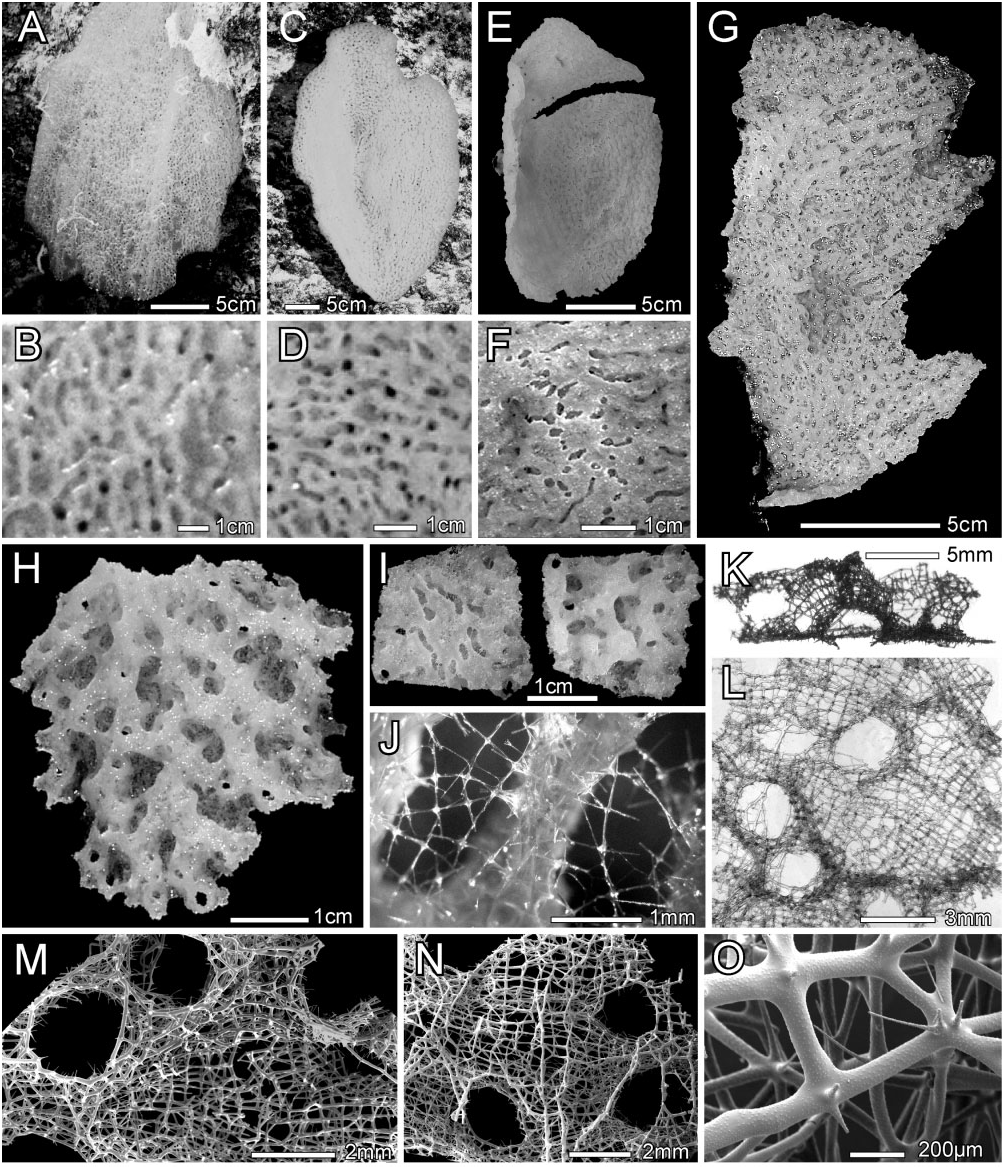
Description: All known specimens have the body form of a thin-walled platter ( Fig. 6A, C, E View Figure 6 ), with corrugations of both dermal and atrial surfaces easily visible in both in situ and deck images ( Fig. 6B, D, F View Figure 6 ). All but one specimen were attached to hard substrate by a short stalk emanating from the centre of the convex (dermal) surface ( Fig. 6E View Figure 6 ); in situ figures indicate that the exception, paratype 1, USNM 1231338, may have been attached marginally ( Fig. 6A View Figure 6 ). Body diameters of paratypes before collection were 41 and 27 cm in diameter; in situ images of the holotype are unavailable, but at the time of the encounter, the experienced observer estimated its diameter as 30 cm and the fragment collected from it was 16.6 × 9.4 cm ( Fig. 6G View Figure 6 ). Corrugations seen in images are the lateral walls of a two-dimensional system of conjoined branching and meandering tubules seen on the dermal side as a system of inflated ridges (the tubules), which circumscribe a variety of isolated irregular depressions ( Fig. 6H View Figure 6 ). The tubules all share a common wall, the atrial wall of the whole specimen, and are thus never free-standing cylinders. That atrial surface is comparatively flat. Small oscula, 0.9–1.7– 2.5 mm diameter ( n = 53), occur through the whole wall marginally, or through the inflated upper wall of ridges on the dermal side and through the atrial wall under those ridges on the atrial side. Total thickness of the holotype is 1.9–3.5– 4.7 mm ( n = 15) and of paratype 1 is 0.3–2.2– 4.2 mm ( n = 51). Dermal ridges of the holotype ( Fig. 6H View Figure 6 ) are 1.5–5.4– 11.1 mm long ( n = 39) and 0.8–2.5– 5.2 mm wide ( n = 66). Grooves of the atrial surface that underlie the dermal ridges are 1.4–3.6– 10.2 mm long ( n = 48) by 0.7–1.1– 1.5 mm wide ( n = 48); medially they are covered by a loose spicule lattice ( Fig. 6J View Figure 6 ). The texture is hard but slightly flexible, brittle, and crumbly; color in situ is pale green but changes to olive green after collection on deck and preservation in ethanol. No symbionts were noted. Distribution as presently known is restricted to waters around San Salvador, Bahamas ( Fig. 1 View Figure 1 ).
The fused skeleton varies in thickness and complexity from marginal to medial areas of the discoid body; dimensions of elements are summarized in Table 3. Marginally, the framework consists of a two-dimensional plate with very indistinct ridges on the dermal side and grooves underlying these on the atrial side. In more medial areas the dermal ridges gradually increase in height by the formation of distinct lateral walls, and the atrial grooves thus become deeper but remain en- tirely open. More medially, the ridges increase in height and the atrial grooves become narrower by encroachment of dictyonal walls; the grooves are now covered by a loose spicule lattice and the tubules circumscribed by them have nearly cylindrical lumina. In the most mature areas the atrial grooves become entirely covered by dictyonal lattice and the tubules within the wall are completely encased in dictyonal framework. The fused skeleton ( Fig. 6K–O View Figure 6 ) of the mature stage consists of two distinct components: (1) thin sheets of somewhat regular rectangular and triangular meshes and (2) thick strands mainly developed in the atrial wall surface ( Fig. 6N View Figure 6 ). The thin sheets form the primary framework of the dermal surface, as the sides and roof of the dermal ridges ( Fig. 6M View Figure 6 ), the floor of the dermal pits, and the cover over the atrial grooves. Meshes have slightly longer longitudinal beams that are aligned over short distances of up to seven meshes to form poorly recognizable longitudinal strands. The sheets are between one and three meshes in thickness, and where the sheet is more than one layer in thickness, the nodes and beams of the adjacent layers are not aligned. This framework is clearly not farreoid, euretoid, nor aulocalycoid. The thick strands form borders of both dermal and atrial oscula as well as obliquely lateral strands on the atrial surface, probably forming marginal support for the atrial grooves. Dictyonal walls of the tubules are 0.16–0.44– 1.13 mm ( n = 8) in thickness. All nodes are regular (unswollen); nodes, beams, and spurs are finely and evenly spined ( Fig. 6O View Figure 6 ). Spurs are very abundant throughout the framework; they are long, thin, and finely pointed on the regular sheet part, but shorter, thicker, and slightly swollen terminally on the thick beams. There are no small oxyhexactins appended to beams.
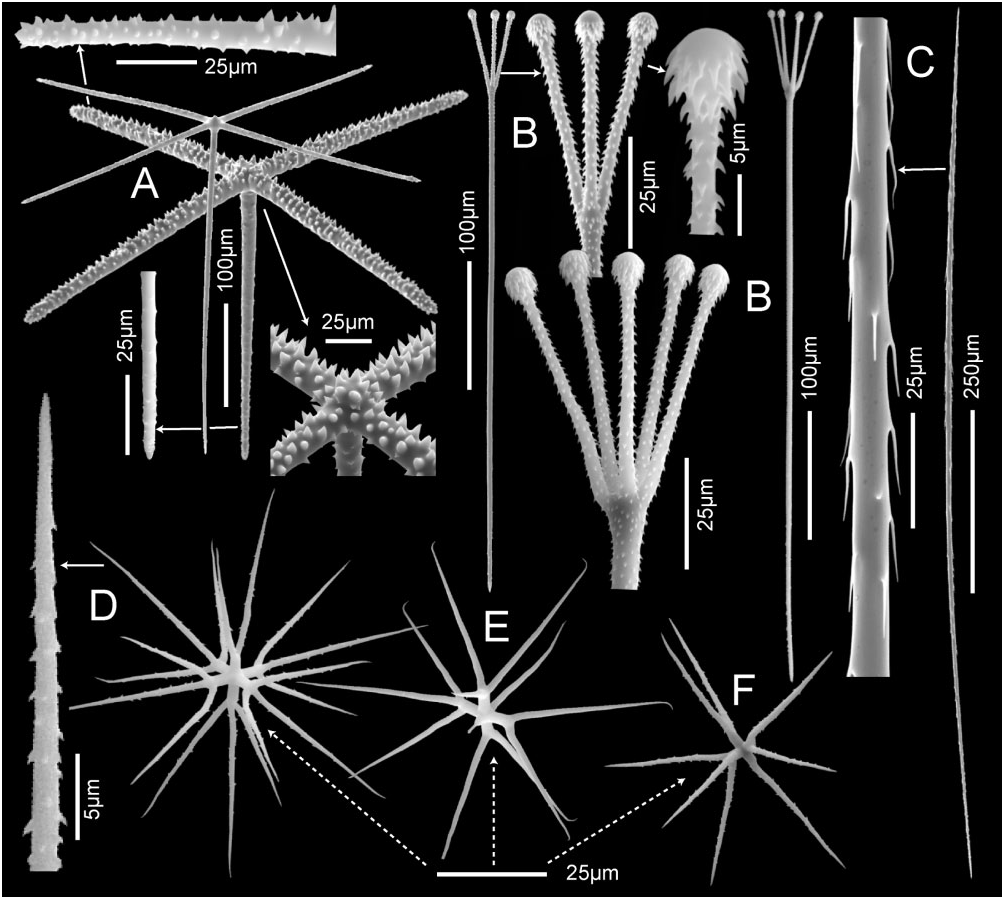
Megascleres consist of surficial pentactins, associated tyloscopules, and uncinates; spicule dimensions are summarized in Table 3. Pentactins ( Fig. 7A View Figure 7 ) vary in thickness; the more robust forms have large conical spines densely covering the outer surfaces of tangential rays; proximal rays of the robust forms and all rays of the thin forms have a dense cover of small spines. These spicules form a quadrate lattice on the dermal and atrial surfaces as well as the internal lining of the tubules. Dermal and atrial pentactins have similar tangential rays, means 271 × 13.6 and 274 × 13.1 μm respectively, but differ in atrial pentactins having slightly longer proximal rays, 260 × 10.6 μm versus 311 × 11.8 μm, respectively. Tyloscopules, mean total length of 514 μm ( Fig. 7B View Figure 7 ), vary considerably in the size of terminal swellings; their small heads bear between two and five straight tines that are moderately spread and covered with reclined spines. Abnormally formed heads with tines lat- erally offset are common. Shafts are either entirely rough or mostly smooth, but always spined at their basal ends; the shaft tip is simple and abruptly pointed without inflation. Tyloscopules are distributed as the pentactins, and usually have the entire head project above surfaces. Uncinates ( Fig. 7C View Figure 7 ) are moderate in size, have a mean length of 1351 μm, with well-developed brackets, and barbs only slightly inclined from the spicule surface. They are present throughout the skeleton, oriented obliquely and radially, and project from all surfaces.
Microscleres consist mostly of regular oxyhexasters (65%), with fewer hemioxyhexasters (19%) and irregular variants (16%); no oxyhexactins are present. Regular oxyhexasters, with mean diameter of 80 μm ( Fig. 7D View Figure 7 ), have short mostly smooth primary rays of 5.1 μm in length, that carry between two and four long, straight, basally and/or distally hooked terminal rays, 35.6 μm in length, liberally covered with reclined spines. Hemioxyhexasters are similar, but one or two primary rays carry only a single terminal ray. Common irregular variants lack development of one or more primary rays, and include oxy- and hemioxypentasters ( Fig. 7E View Figure 7 ), oxy- and hemioxytetrasters ( Fig. 7F View Figure 7 ), oxy- and hemioxytriasters, oxydiasters and oxyspirasters. All microsclere forms are generally distributed.
Etymology: The species name, ‘ lecus ’, is derived from ‘ lekos ’, Gk for disc, with reference to the body form of this new species.
Remarks: The presence of a dictyonal framework and scopules mandates assignment of this species to suborder Sceptrulophora . Absence of framework channellization prevents its assignment to families Aphrocallistidae , Craticulariidae , Cribrospongiidae , or Tretodictyidae . Lack of a two-dimensional farreoid framework prevents its inclusion in Farreidae . Presence of two distinct types of framework, one with thickened beams, prohibits assignment to Euretidae and Fieldingiidae , but is consistent only with its placement in Auloplacidae . Its basic structure of branching tubular elements sharing a common wall and spiculation are also consistent with that assignment. It cannot conveniently be accepted as a member of the genus Auloplax because the pattern of constituent tube origin and growth differ from that known in all Auloplax species. In Auloplax , tubes branch from a basal tube, rebranch several times, but always remain joined to their last branched tube by shared common walls; the tubes may be partly free terminally but never anastomose. In the new species, the tubes never share common lateral walls but they all indirectly share a common atrial wall; the tubes divide and anastomose, forming a network of tubes not seen in any member of Auloplax . All three or four species of Auloplax [ Auloplax filholi ( Topsent, 1904) has not yet been definitely shown to be a junior synonym of A. auricularis Schulze, 1904 ] have discohexactins or discohexasters as their major microsclere, whereas the new species has regular oxyhexasters instead. There is a precedent for Auloplacidae with oxy-microscleres in the form described as Auloplacidae incertae sedis in Reiswig & Kelly (2011). The differences in body form, constituent tube growth, and microsclere form from all Auloplax species mandates the erection of a new genus for the new species, proposed here as Dictyoplax lecus sp. nov.

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |