
Apleurotropis Girault
|
publication ID |
https://doi.org/ 10.5281/zenodo.197172 |
|
DOI |
https://doi.org/10.5281/zenodo.6210827 |
|
persistent identifier |
https://treatment.plazi.org/id/03B3A716-FFD9-3919-FF4D-B2DF46C3AAB8 |
|
treatment provided by |
Plazi |
|
scientific name |
Apleurotropis Girault |
| status |
|
Apleurotropis Girault View in CoL View at ENA
Apleurotropis Girault, 1913: 77 View in CoL –78. Type species: Apleurotropis viridis Girault View in CoL , by original designation.
Apleurotropopseus Girault, 1913: 146 . Type species: Apleurotropopseus albipes Girault , by original designation. Synonymized by Bouċek, 1988: 711.
Epentedon Girault, 1915: 186 . Type species: Epentedon unnotipennis Girault , by original designation. Synonymized by Bouċek, 1988: 711.
Horismenella Girault, 1915:189 . Type species: Horismenella clariviridis Girault , by original designation. Synonymized by Bouċek, 1988: 711.
Pleurotropomyia Girault, 1913: 145 . Type species: Pleurotropomyia grotiusi Girault , by original designation. Synonymized by Bouċek, 1988: 711.
Propleurotropis Girault, 1937:1 –2. Type species: Propleurotropis minii Girault , by original designation. Synonymized by Bouċek, 1988: 711.
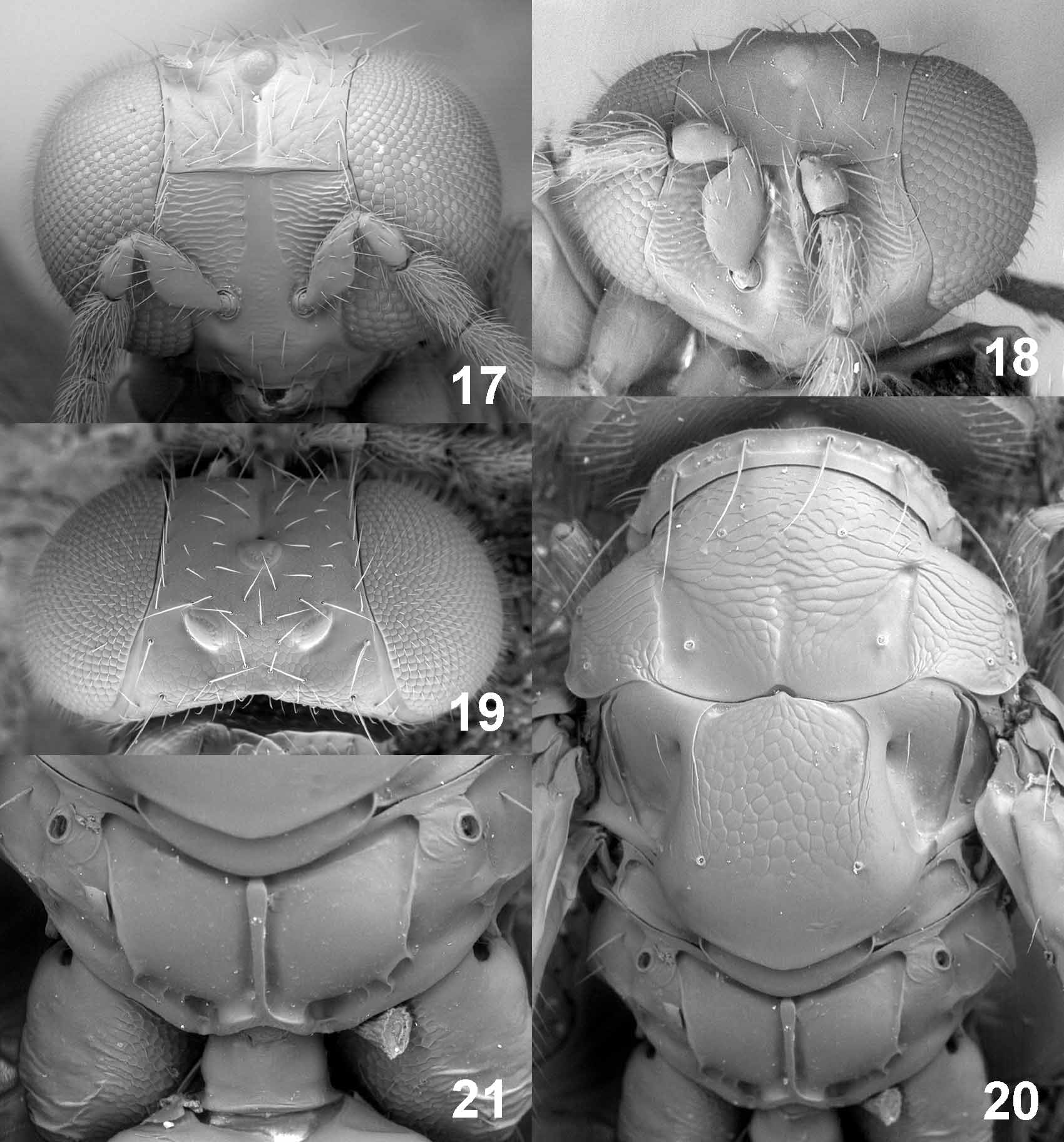
Diagnosis. Eyes with dense long hairs (e.g. Figs 7 View FIGURES 7 – 11 , 12 View FIGURES 12 – 16 ); vertex with a pit or a short groove in front of anterior ocellus (e.g. Figs 4 View FIGURES 2 – 6 , 9 View FIGURES 7 – 11 ); frons just above frontal suture raised to form a carina, V-shaped, straight or slightly down-curved laterally (e.g. Figs 2 View FIGURES 2 – 6 , 17 View FIGURES 17 – 21 , 22 View FIGURES 22 – 26 ); pronotum with a transverse carina close to posterior margin (e.g. Figs 5 View FIGURES 2 – 6 , 10 View FIGURES 7 – 11 ), laterally also with a complete longitudinal carina ( Fig. 31 View FIGURES 27 – 31. 27 – 30 ); mesoscutum usually with a median groove in posterior 1/3–1/2 (e.g. Figs 5 View FIGURES 2 – 6 , 15 View FIGURES 12 – 16 ); mesoscutum and scutellum with engraved and strong reticulation (e.g. Figs 10 View FIGURES 7 – 11 , 20 View FIGURES 17 – 21 ); postmarginal vein long, 2.4–3.7X as long as stigmal vein; propodeum with complete and more or less parallel plicae and with a complete median carina (e.g. Figs 6 View FIGURES 2 – 6 , 11 View FIGURES 7 – 11 , 16 View FIGURES 12 – 16 ).
Description. Flagellum with a two-segmented clava in both sexes; male flagellomeres with scattered setae; male scape enlarged, ventral sense area present along entire scape; sensilla ampullacea symmetric and short, present on all flagellomeres. Antenna with discoid anelli. Mandibles with two large teeth at apex and with one small tooth above large teeth. Clypeus not delimited. Malar sulcus present. Upper frons just above frontal suture as a raised carina, straight or slightly down-curved laterally; antennal scrobes reach frontal suture separately. Occipital margin rounded; occiput without median groove or fold above foramen magnum.
Pronotum with a transverse carina close to posterior margin. Midlobe of mesoscutum with two pairs of setae; mesoscutum and scutellum with engraved reticulation; mesoscutum with a shallow median groove in posterior 1/3–1/2; notauli distinct in anterior ½, in posterior ½ present as weakly delimited notaular depressions that are usually smooth. Scutellum with or without lateral grooves in anterior 1/2–2/3; with one pair of setae. Transepimeral sulcus weakly curved. Dorsellum visible in dorsal view. Postmarginal vein 2.4– 3.7X as long as stigmal vein. Propodeum smooth and shiny with complete median carina and parallel and complete plicae.
Petiole 0.5–2.0X as long as wide, smooth and shiny or with irregular carinae, with or without a spine medially to either side of the petiole, ventral surface smooth. The male genitalia are similar to the majority of Entedoninae , i.e. with two digital spines and with volsellar setae thin and narrow (see e.g. Hansson 1996).
Biology. Primary parasitoids on leaf-mining larvae of sawflies ( Hymenoptera ), Agromyzidae (Diptera) , Hispinae ( Coleoptera , Chrysomelidae ), and various Lepidoptera groups ( Bouček 1988). Hosts for the species described here are mainly from leaf-mining Lepidoptera , with one record from a leaf-mining Diptera and one from an unidentified gall (all new records): Bucculatrix sp. ( Lepidoptera : Bucculatricidae ) on Forsteronia spicata (Apocyanaceae) ; Elachista sp. ( Lepidoptera : Elachistidae ) on Olyra latifolia (Poaceae) ; Marmara sp. ( Lepidoptera : Gracillariidae ) on Heliconia sp. ( Heliconiaceae ); Phyllocnistis sp. ( Lepidoptera : Gracillariidae ) on Trichilia havanensis (Meliaceae) ; Tischeria spp. ( Lepidoptera : Tischeriidae ) on Coussapoa villosa (Moraceae) and Gouania polygama (Rhamnaceae) ; unidentified Elachistidae (Lepidoptera) mining leaves of Bambusa (= Guadua ) angustifolia (Poaceae) ; unidentified lepidopteran leafminers on Saurauia sp. ( Actinidiaceae ), Trichilia havanensis , Peperomia peltata (Piperaceae) , unidentified Asteraceae , Heliconiaceae and Melastomataceae ; Agromyzidae indet. ( Diptera ) on Tradescantia sp. ( Commelinaceae ); from a leaf pustule gall on Ficus hartwegii (Moraceae) .
Distribution. Australia, India, Japan, Madagascar, New Zealand, Papua New Guinea, Tanzania. Tropical America (all new records): Brazil, Colombia, Costa Rica, Dominican Republic, Ecuador, Honduras, Mexico, Peru.
Remarks. Apleurotropis shares some characters with Achrysocharoides Girault : hairy eyes, frons just above frontal suture raised and forming a carina, lateral downsloping part of pronotum with a longitudinal carina. This latter character is somewhat different in the two genera, in Achrysocharoides it is incomplete, present only in posterior ½, whereas it is complete in Apleurotropis . The two latter characters are as far as is known present only in these two genera, hence indicating a possible relationship. These genera differ as follows. Apleurotropis has antennal scrobes in females joining frontal suture separately, mesoscutum and scutellum with engraved reticulation, postmarginal vein 2.4–3.7X as long as stigmal vein, propodeum with a distinct and strong median carina; Achrysocharoides with antennal scrobes in females joining below frontal suture, mesoscutum and scutellum with raised reticulation or partly smooth — sometimes with pits, postmarginal vein 0.5–1.5X (usually about 1X) as long as stigmal vein, and propodeum without strong median carina.
Identification. In the key to the Nearctic genera of Eulophidae by Schauff et al. (1997) Apleurotropis runs to couplet 119 where the genera Entedon and Pediobius are keyed out. Apleurotropis differs from both in having engraved reticulation on midlobe of mesoscutum and scutellum (mesoscutum and scutellum with raised reticulation in both Entedon and Pediobius ), and in having a very long postmarginal vein (2.4–3.7X as long as stigmal vein), in Entedon and Pediobius the postmarginal vein is at most 1.7X as long as stigmal vein but usually these two veins are subequal in length. It differs from Entedon in having distinct propodeal plicae and from Pediobius in having medioposterior part of propodeum short and anterior part of petiole narrow — in Pediobius the medioposterior part of propodeum is extended backwards to varying degrees to form a nucha, and anterior part of petiole is wide and concave to embrace the extended medioposterior part of propodeum. Another option for identification is to use the multiple entry key to the Neotropical genera of Entedoninae on the website www.neotropicaleulophidae.com.





No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
Apleurotropis Girault
| Hansson, Christer 2010 |
Propleurotropis
| Boucek 1988: 711 |
| Girault 1937: 1 |
Epentedon
| Boucek 1988: 711 |
| Girault 1915: 186 |
Horismenella
| Boucek 1988: 711 |
| Girault 1915: 189 |
Apleurotropis
| Girault 1913: 77 |
Apleurotropopseus
| Boucek 1988: 711 |
| Girault 1913: 146 |
Pleurotropomyia
| Boucek 1988: 711 |
| Girault 1913: 145 |