
Ampliphax grandis, Bartlett, Charles R. & Kunz, Gernot, 2015
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3946.4.2 |
|
publication LSID |
lsid:zoobank.org:pub:5ACF3615-12A9-46FA-8950-4506692DCDEB |
|
DOI |
https://doi.org/10.5281/zenodo.6108064 |
|
persistent identifier |
https://treatment.plazi.org/id/03B30A49-FFB3-7E75-FF6F-1D8D55D9D3D3 |
|
treatment provided by |
Plazi |
|
scientific name |
Ampliphax grandis |
| status |
sp. nov. |
Ampliphax grandis View in CoL new species
( Figures 1–4 View FIGURE 1 View FIGURE 2 View FIGURE 3 View FIGURE 4 )
Type locality. Costa Rica, Puntarenas Province, Estacion Pittier, near Cerro Gemelo, 1670m
Diagnosis. A large, orange-stramineous species with a projected head, keeled lateral carinae of the vertex and terete antennae. Calcar large, foliaceous, with greater than 25 small, black-tipped teeth on the trailing margin. Male genitalia with simple parameres, a simple decurved aedeagus, and very large curved pair of appendages on the genital diaphragm.
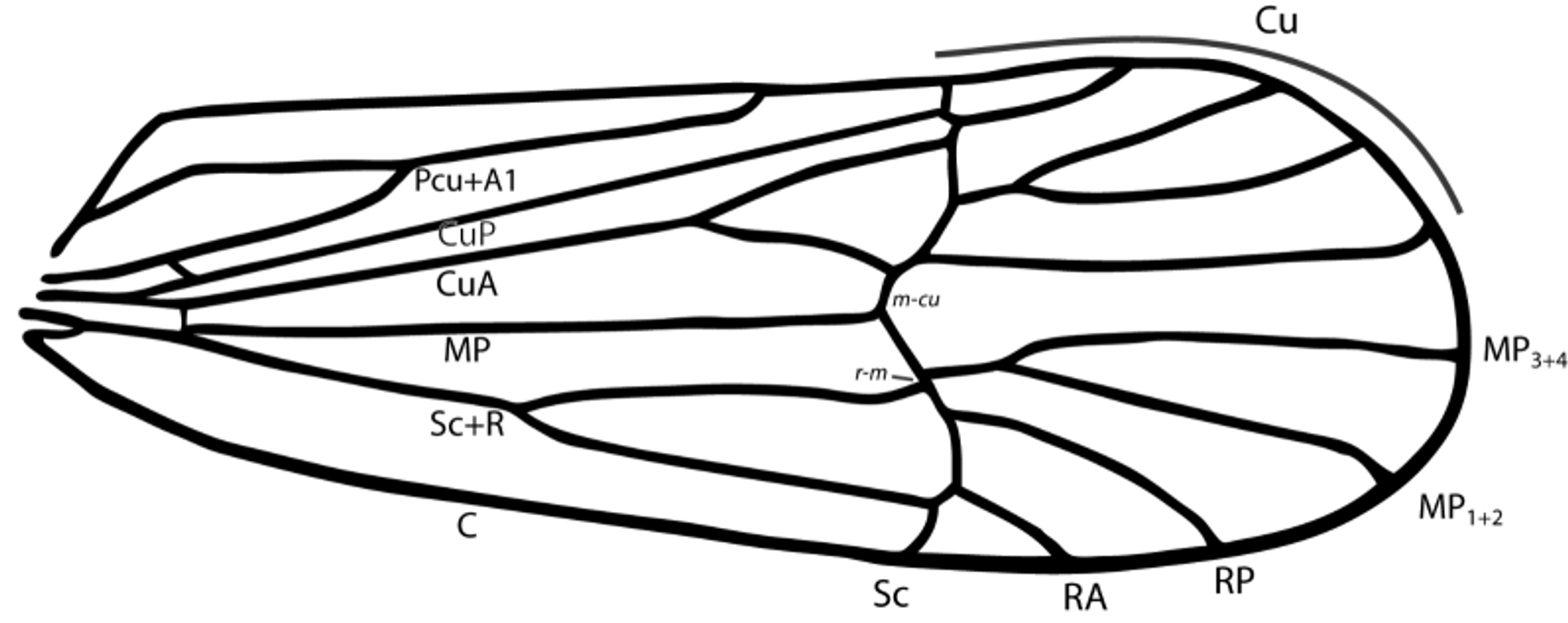
Description. Color. Uniformly orange-stramineous to orange-tan (some specimens weakly washed anteriorly with grey), paler in median regions of vertex, pro- and mesonotum. Macropterous wings clear to weakly infuscate. Structure. Macropter. Body length, male, without wings x = 3.11 mm (range 3.03–3.20, n=3), with wings x = 5.19 mm (range 5.14–5.27, n=4), forewings 4.12 mm (range 4.02–4.21, n=3); female (without wings) x = 3.80 (range 3.61–4.12, n=4), with wings 5.78 (range 5.67–5.85, n=4). Head. Head narrower than pronotum. Vertex longer than wide (l: w 1.69:1, n=5). Head narrower than pronotum, in lateral view distinctly anteriorly projecting, appearing slightly upcurved. Carinae of head and thorax conspicuous, except Y-shaped and submedian carinae of vertex obsolete; lateral margins of vertex strongly carinate, meeting at fastigium. Frons rather parallel-sided about 3 × longer than wide (w:l 0.27:1), widest near ventral margin of eyes, narrowed between eyes; carinae sharp, median carina of frons narrowly forked near fastigium. Antennal segment I about twice as long as wide, antennal segment II about 2 × longer than I (ratio I:II 0.45:1) Thorax. Prothorax about 1/3 length of mesothorax (length ratio 0.37:1); carinae evident, lateral carinae diverging, not reaching posterior margin. Mesonotum with carinae evident, reaching posterior margin, lateral carinae slightly diverging posteriorly. Forewing venation varying in details among individuals. Forewing with nodal line in distal third; Sc+R arising from leading margin of basal cell, forked into Sc+RA and RP near midlength of clavus, Sc forked from RA at nodal line, RA unbranched; RP unbranched or forked following nodal line; MP forked from SC+R+M (+CuA according to Nel et al., 2012, Bourgoin et al. 2015) at basal cell, abruptly angled at nodal line and subsequently to MP1+2 and MP3+4 at (or after) nodal line, some specimens with MP3+4 subsequently branched; CuA arising from trailing margin of basal cell, forked into CuA1 and CuA2 distad of Sc+RA and RP fork; with CuA1 branch often subsequently forked. Calcar large, flattened and foliaceous, narrowed distally, bearing continuous row of more than 28–35 black-tipped teeth on posterior lateral margin, subequal or exceeding length of metabasitarsus. Metabasitarsus slightly more than half length of tarsus (0.55:1, n=5). Abdomen. Pygofer in lateral view quadrate, taller than wide in lateral view; anterior margin truncate, caudal margin sinuate; ventrocaudal margin with elongate, scooplike projection. In caudal view, pygofer opening keeled, taller than wide; ventral margin of opening with prominent projection, ventrolateral margins strongly keeled. Diaphragm deeply emarginate medially, armature consisting of a pair of very large caudally declined scalpriform appendages arising from thickened base; avicephaliform in lateral view, U-shaped in caudal view. Parameres narrow, flattened, parallel-sided, basal angles obscure; diverging in basal two-thirds, angled dorsal in apical third, apex abruptly angled lateral; apex truncate; in lateral view somewhat dorsocaudally directed. Aedeagus simple, decurved, parallel-sided and weakly flattened, bearing small subapical lateral flanges of few teeth. Abdominal segment X quadrate, bearing short, thick sharply-pointed processes on dorsocaudal angles; widely separated at base in caudal view; segment XI subequal to height of segment X.
Remarks. Given its relatively large size and distinctive features, relatively few specimens of Ampliphax grandis have been found. Of the 32 specimens of this species recorded here, the sex ratio was highly biased toward females, with 26 females and 6 males. The few specimens with reported collection methods came from lights, although some specimens collected by F. S. Blanton had debris on them suggesting they were obtained from Malaise traps. No host plants have been recorded.
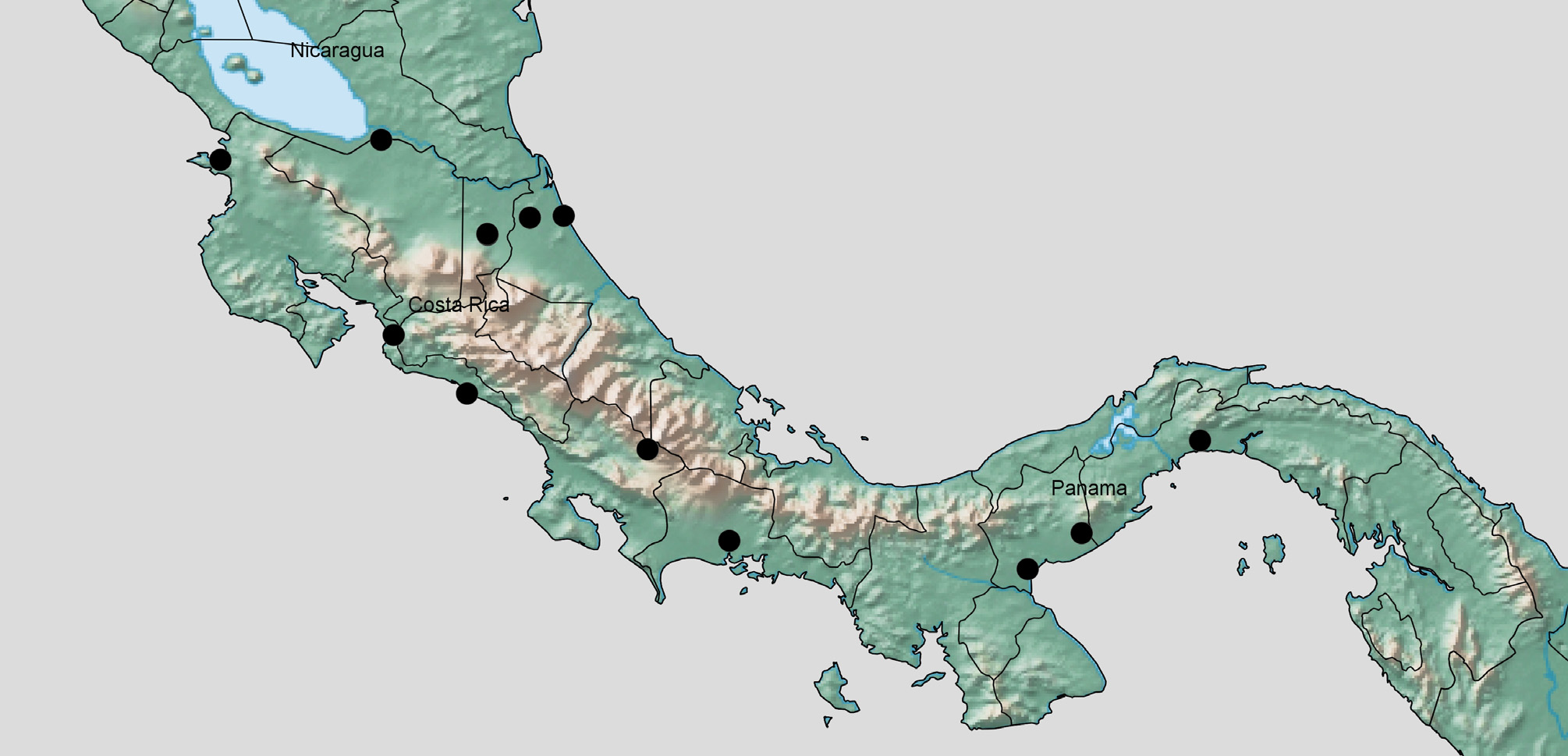
A general zoogeographic pattern found in Costa Rica has been that different, but closely related, species are found among the Pacific tropical dry forest (lowland) region, the higher elevations (especially the Talamanca Mountains), and the Atlantic lowlands. We considered this pattern, but available evidence suggests a single species. However, the available evidence is limited because all the specimens from Guanacaste and Alajuela provinces, Costa Rica (in the northwest), were female, as were available specimens from Panama. The genitalia of the 6 males (from Limon, Puntarenas, and Heredia provinces, Costa Rica) were similar. The most notable differences were that the holotype (from Puntarenas, 1670 m), had a more pointed aedeagus and longer processes on segment X than the specimen illustrated (Heredia, ~ 60 m). Also, DNA barcodes show that the holotype has less than 1% difference from a specimen from Limon (10 m elevation).
A single female specimen from the Dominican Republic was excluded from the paratype series because of its disjunct range. A specimen from Laguna Medio Queso, Costa Rica was excluded because it was in poor condition.
The holotype has been DNA barcoded with the full 658 bp sequence available on BOLD (process id ASIHE1471–12, BIN URI ABZ9263).
Reported hosts. None.
Distribution. Costa Rica, Panama ( Fig. 4 View FIGURE 4 ); also tentatively from Dominican Republic.
Etymology. The specific epithet is from the Latin word grandis (large, great, magnificent), and has a common termination in masculine and feminine form.
Type material examined. HOLOTYPE (male, INBio). COSTA RICA. Puntarenas, Est. / Pittier, 4.2 Km SO. del Cerro / Gemelo 1670m, 8–20 JUL 1997. / M. M. Moreaga. Red de Golpe / L_S_330900_577400 #47390 // [inverted barcode label] COSTA RICA / INBio / CRI 002 / 567442 // DNA Barcoding / E. Ulate / CCDB –11593 D10 // HOLOTYPE / Ampliphax / grandis / Bartlett & Kunz [red paper].”
PARATYPES. COSTA RICA: Guanacaste: Rio Pedregal, Murcielago, A.C.G., P.N. Santa Rosa, 100 m, Nov 1993, F. Munoz, L N [Lambert north] 320300 347200 #2506 (4f INBio); Rincón del Vieja, National Park Rincón de la Vieja, N10°45´15´´; W85°21´00´´, 635m, 10 June 2008, G. Kunz (1f, photographs, Fig. 1 View FIGURE 1 ); Heredia: La Selva, 50 m, 0 1 Sep 1998, C. W. & L. B. O'Brien, at light (1f, LBOB). near Puerto Viejo, La Selva Biological Station, 10.41667 ° N 84 ° W, 55 m, 17 Aug 2003, C.R Bartlett, J. Cryan and J. Urban (1f, UDCC); same, 23 Feb 2004 – 02 Mar 2004 (1f, UDCC); same, 25–26 Feb 2004, C. R. Bartlett (1f, UDCC); same, 29 Feb 2004, C. R. Bartlett, mercury vapor light, Voucher NCSM tissue collection 09–11–20–35 (1m, UDCC, 1f, NCSM, in alcohol); Limon: Est[acion] Cuatro Esquinas, P. N. Tortuguero, Jun 1990, E. Quesada, L N 280000, 590500 (1f, INBio); Sector Cedrales de la Rita, 3 km N del Puente Rio Suerte, Ruta Puerto Lindo, 10 m, Jan 1997, E. Rojas, L N 278600, 566500 (2f, INBio, CRI 002545535 DNA barcoded); same, Feb 1997 (1f, 2m INBio); same, Apr 1997 (3f, 1m INBio); Puntarenas: Estacion Pittier, 4.2 Km SW del Cerro Gemelo, 1670 m, 08–20 Jul 1997, M. M. Moreaga, Red de Golpe, L S [Lambert south] 330900, 577400 (1m, INBio); Estacion Quebrada Bonita, Res. Biol. Carara, 50 m, May 1990, R. Zuniga, [L S?] 194500, 469850 (1f, INBio); Quepos, P.N. Manuel Antonio, 80 m, Feb 1991, R. Zuniga, L S 370900, 448800 (1f, INBio, CRI 000312790 barcoded). PANAMA: Chiriqui: David, Oct 1959, N. L. H. Krauss (1f, USNM); Cocle: El Retiro, 10 Nov 1952, F. S. Blanton (1f, USNM); Pt. [Port of] Aguadulce, 21 Nov 1952, F. S. Blanton (1f, USNM); Panama: Tocumen, 20 Dec 1951, F. S. Blanton (3f, USNM).
Other material examined. COSTA RICA: Alajuela: Laguna Medio Queso, 0–100 m, 0 1 Sep 2005, Y. Cardenas, J. Azofeifa, M. Moraga, Red Noyes, L N [Lambert north] 334350, 461100, #84534 (1f, INBio). DOMINICAN REPUBLIC: San Cristóbal Province, San Cristóbal, July 1969, J. Maldonado C. (1f, USNM) (excluded from paratypes).
Molecular data. Three specimens of this species from Costa Rica (viz. CRI 000312790, CRI 002567442, CRI 002545535) have been DNA barcoded by INBio (a 648 base-pair region in the mitochondrial cytochrome c oxidase 1 gene) with data reported in the Barcode of Life Database ( BOLD, www.boldsystems.org; Ratnasingham & Hebert 2007, 2013) as part of the Delphacidae Costa Rica INBio Collection ( DELCR) project. The sequences differ from each other with a p-distance of less than 1%. The nearest neighbor in BOLD is Bostaera balli with a pdistance statistic given at 13.96%, indicating that no taxon closely related to Ampliphax has been barcoded. Pareuidella and ‘ Euides ’ were among the taxa with barcode data in BOLD (but not Neoperkinsiella among specimens with determinations). BOLD has barcode data for 1,261 specimens, barcodes representing 287 species, in comparison to approximately 576 currently described New World delphacid species (2100+ species worldwide, Bourgoin 2014), suggesting that the closest relatives to Ampliphax may not be barcoded to date.


| INBio |
National Biodiversity Institute, Costa Rica |
| CRI |
Universidade do Extremo Sul Catarinense, Bairro Universitário |
| DNA |
Department of Natural Resources, Environment, The Arts and Sport |
| CCDB |
Crustacean Collection of the Department of Biology |
| UDCC |
University of Delaware |
| NCSM |
North Carolina Museum of Natural Sciences |
| USNM |
Smithsonian Institution, National Museum of Natural History |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |