
Leporinus torrenticola, Birindelli, José L. O., Teixeira, Tulio F. & Britski, Heraldo A., 2016
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4178.1.4 |
|
publication LSID |
lsid:zoobank.org:pub:0C795CEE-65CE-4903-9B64-DBC18BFC0A06 |
|
DOI |
https://doi.org/10.5281/zenodo.5624760 |
|
persistent identifier |
https://treatment.plazi.org/id/03B25219-9B3C-FFFF-FF31-8DE6629D922D |
|
treatment provided by |
Plazi |
|
scientific name |
Leporinus torrenticola |
| status |
sp. nov. |
Leporinus torrenticola , new species
( Figs. 5–7 View FIGURE 5 View FIGURE 6 View FIGURE 7 )
Leporinus View in CoL sp.— Zuanon, 1999: 87, figs. 3.1, 3.12 (diagnosis, morphometric data for ecomorphological analysis, behavior, feeding, picture of live specimen, illustrations of head and teeth; Pará, Altamira, rio Xingu).
Holotype. MNRJ 34071, 1 , 92.9 mm SL; Brazil: Pará: Altamira: Xingu basin: rio Curuá , tributary of rio Iriri , at Castelo dos Sonhos, 8°19'06"S 55°05'24"W; J. L. O. Birindelli, M. R. Britto & J. Gomes, 30 Sep 2008. GoogleMaps
Paratypes. Brazil, Pará (Xingu basin). ANSP 201879, 2 , 94.4–119.6 mm SL; Altamira: rio Curuá, rio Iriri basin, downstream large waterfalls, 8°43'50"S 54°57'49"W; J. L. Birindelli et al., 20 Oct 2007. GoogleMaps — INPA 53126, 2 , 101.5– 112.1 mm SL; same data as ANSP 201879 About ANSP . GoogleMaps — MZUEL 14500, 2 , 95.9–108.1 mm SL; same data as ANSP 201879 About ANSP . GoogleMaps — MNRJ 47512, 2 , 103.8– 116.9 mm SL; same data as ANSP 201879 About ANSP . GoogleMaps — MZUSP 96950, 1 , 90.6 mm SL; Altamira: rio 13 de Maio, tributary of rio Curuá, rio Iriri basin, under the bridge of road BR 163, nearby the village of Cachoeira da Serra , 8°38'53"S 55°01'41"W; J. L. Birindelli et al., 20 Oct 2007. GoogleMaps — MZUSP 97132, 1 , 83.6 mm SL; Altamira: rio 13 de Maio, tributary of rio Curuá, rio Iriri basin, downstream Cachoeira Grande , 8°43'41"S 55°01'38"W; J. L. Birindelli et al., 22 Oct 2007. GoogleMaps — MZUSP 97156, 8 , 74.7–114.4 mm SL, 1 CS, 93.2 mm SL; Altamira: rio Curuá, rio Iriri basin, village of Castelo dos Sonhos, 8°19'07"S 55°05'23"W; J. L. Birindelli et al., 22 Oct 2007. GoogleMaps — MZUSP 96967, 7 , 95.7–123.7 mm SL; same data as ANSP 201879 About ANSP . GoogleMaps
Non-type specimens. Brazil, Pará (Xingu basin). ANSP 193014, 5 , 74.2–120.7 mm SL; Altamira: rio Iriri at Cachoeira Grande , 3°50'36"S 52°44'08"W; M. H. Sabaj Pérez et al, 10 Oct 2012. GoogleMaps — ANSP 194751, 6 , 78.2–140.6 mm SL; Anapu: rio Xingu, 1 km above Cachoeira de Jericoá , 3°21'56"S 51°43'36"W; M. H. Sabaj Pérez et al., 14 Sep 2013. GoogleMaps — ANSP 194809, 5 , 59.0– 71.1 mm SL; Altamira: rio Xingu, 14 km above mouth of rio Iriri , 3°56'21"S 52°34'44"W; M. H. Sabaj Pérez et al., 10 Sep 2013. GoogleMaps — ANSP 194410, 1 , 126.9 mm SL; Altamira: rio Xingu , 3°16'41"S 52°02'18"W; M. H. Sabaj Pérez et al., 18 Sep 2013. GoogleMaps — ANSP 196916, 1 , 35.6 mm SL; Altamira: rio Iriri, c. 14.5 km W-SW of confluence with rio Xingu , 3°51'19"S 52°43'43.5"W GoogleMaps ; M. H. Sabaj Pérez et al.— ANSP 197380, 1 , 95.5 mm SL; Altamira: rio Iriri at Cachoeira Grande , c. 15 km upstream from confluence of rio Xingu , 3°50'32.3"S 52°44'3.9"W GoogleMaps ; M. H. Sabaj Pérez et al.— INPA 9505, 1 , 104.1 mm SL; Altamira: rio Xingu at Cachoeira do Cotovelo , 3°12'43"S 52°12'12"W; M. Jégu, 14 Oct 1992. GoogleMaps — INPA 13191, 3 , 86.6–96.4 mm SL; Altamira: rio Xingu at Costa Júnior , 3°29'28"S 52°19'07"W; J. A. Zuanon, 22 Sep 1997. GoogleMaps — INPA 31011, 7 , 76.9– 92.2 mm SL; Altamira: rio Xingu at Cachoeira do Espelho , 3°39'05"S 52°22'43"W; H. López-Fernandez et al., 24 Aug 2008. GoogleMaps — INPA 38041, 2 , 61.7–76.5 mm SL; same data as ANSP 193014 About ANSP . GoogleMaps — INPA 40077, 1 , 84.5 mm SL; Altamira: rio Xingu, 14 km above mouth of rio Iriri , 3°56'21"S 52°34'44"W; M. H. Sabaj Pérez et al., 10 Sep 2013. GoogleMaps — INPA 40082, 2 , 63.5–104.1 mm SL; same data as ANSP 194809 About ANSP . GoogleMaps — INPA 40204, 8 , 76.0– 104.1 mm SL; same data as ANPS 194751 . GoogleMaps — INPA 40417, 1 , 87.5 mm SL; same data as ANSP 194410 About ANSP . GoogleMaps — INPA 47122, 1 , 73.8 mm SL; Altamira: rio Bacajaí, tributary of the right margin of rio Xingu at Volta Grande; M. H. Sabaj et al., 11 Sep 2014. — LIA 1119, 1 , 64.7 mm SL; São Felix do Xingu: Xadai, upstream from São Felix do Xingu ; L. M. Sousa et al., 20 Sep 2014. — LIA 1258, 3 , 58.2–77.6 mm SL; São Felix do Xingu: Xadai, upstream from São Felix do Xingu , 6°52'15"S 52°02'35"W; L. M. Sousa et al., 23 Sep 2014. GoogleMaps — MZUEL 14129, 2 , 75.9–84.2 mm SL; same data as INPA 40204 View Materials . GoogleMaps – MZUSP 29180, 1 , 48.0 mm SL; Belo Monte: Igarapé do Aturiá, tributary of rio Xingu , 3°07'00"S 51°42'00"W; M. Goulding, 28 Sept 1983. GoogleMaps — MZUSP 29181, 1 , 58.9 mm SL; same data of MZUSP 29180 View Materials . GoogleMaps — MZUSP 29183, 33 , 45.8–111.8 mm SL; Belo Monte: Igarapé do Santo Antônio, tributary of rio Xingu , 3°07'00"S 51°42'00"W; M. Goulding, 27 Oct 1983. GoogleMaps — MZUSP 110624, 1 SK, 120.0 mm SL; Altamira: rio Xingu, above Cachoeira Jericoá , 3°21'53"S 51°43'57"W GoogleMaps ; J. L. O. Birindelli et al., 11 Nov 2011.— MZUSP 111281, 1 , 86.2 mm SL; Altamira: rio Iriri at Cachoeira Grande , 3°50'37"S 52°44'02"W GoogleMaps ; O. T. Oyakawa et al., 16 Nov 2011. – MZUSP 111976, 1 , 70.3 mm SL; Altamira: rio Iriri at Cachoeira Grande , 3°50'37"S 52°44'02"W GoogleMaps ; H. Varella, 22 Oct 2011
Diagnosis. Leporinus torrenticola is distinguished from all other anostomids except Hypomasticus julii , H. megalepis , H. pachycheilus , Leporinus gomesi , L. granti , L. multimaculatus , L. nijsseni , and L. santosi by having one dark blotch on midline of anterior portion of the flank (between opercle and pelvic-fin origin) surrounded by five to seven dark blotches (vs. anterior portion of flank with dark longitudinal stripes, or with dark transversal bars, or with dark blotches not forming the aforementioned pattern). Leporinus torrenticola is distinguished from Hypomasticus julii , H. pachycheilus , Leporinus gomesi , L. granti , L. nijsseni , and L. santosi , by having three teeth on premaxilla (vs. four), and subinferior mouth (vs. inferior in Hypomasticus julii and H. pachycheilus , and terminal in Leporinus gomesi , L. granti , L. nijsseni , and L. santosi ); and from H. megalepis by having 37 to 40 scales in the lateral line (vs. 33 to 35), and from L. multimaculatus by having 16 scale rows around caudal peduncle (vs. 12). See discussion for additional notes on species diagnosis.
Description. Morphometric data in Table 2 View TABLE 2 . Body small, relative to congeners. Largest examined specimen 123.2 mm SL. Head and body elongate and moderately compressed. Dorsal profile convex from snout tip to vertical through anterior nostril, gently convex from latter point to supraoccipital spine, gently convex or straight from that point to dorsal-fin origin, straight along dorsal-fin base, approximately straight from dorsal-fin terminus to adipose fin, and slightly or distinctly concave from adipose fin to anteriormost dorsal caudal-fin procurrent ray. Ventral profile slightly concave or relatively straight from tip of lower jaw to vertical through pectoral-fin origin, convex from that point to pelvic-fin origin, straight or slightly concave from latter point to anal-fin origin, straight along anal-fin base, and concave from anal-fin terminus to anteriormost ventral caudal-fin procurrent ray. Greatest body depth at dorsal-fin origin.
Mouth subinferior, its cleft longitudinally aligned with ventral margin of eye in specimens around 70 mm SL, and with ventral margin of infraorbital series in specimens 80 mm SL or larger. Snout rounded. Premaxillary bone with 3* (51) incisiform teeth gently decreasing in size from symphyseal tooth laterally ( Fig. 7 View FIGURE 7 ). Dentary bone with 4* (46) or 5 (1) incisiform teeth also gently decreasing in size laterally. Teeth with two or more rudimentary cusps in specimens of around 60 mm SL or less, cusps more developed in lateralmost teeth.
Scales cycloid, with 9 to 20 (3) radii reaching posterior margin. Lateral line complete with 37 (7), 38 (20), 39* (23) or 40 (2) perforated scales, extending from supracleithrum to base of median caudal fin rays. Horizontal scale rows between dorsal-fin origin and lateral line 4 (2) or 5* (50). Horizontal scale rows between lateral line and pelvic-fin origin 4* (29) or 4.5 (16). Horizontal scale rows around caudal peduncle 15 (1) or 16* (51). Predorsal scales from tip of supraoccipital spine to dorsal-fin origin 11* (20), 12 (8) or 13 (1).
Dorsal-fin rays ii,10* (30); its origin slightly anterior to vertical through pelvic-fin insertion and anterior to middle of standard length; distal margin convex or straight. Adipose fin small, teardrop shaped, its origin through vertical base of fourth or fifth anal-fin ray. Pectoral-fin rays i,13 (1), i,14 (9), i,15* (19) or i,16 (1); tip of fin extending from half to two-thirds distance between of pectoral and pelvic-fin insertions; distal margin straight or convex. Pelvic-fin rays i,8* (30); origin at vertical through base of second to fourth dorsal-fin branched ray; distal margin rounded or straight. Anal-fin rays ii,8* (36); origin slightly posterior to half the distance between pelvic-fin origin and ventralmost caudal-fin procurrent ray; adpressed fin almost reaching anteriormost caudal-fin procurrent rays. Distal margin of anal fin convex in most specimens, with anterior branched ray slightly less than twice last branched ray, or straight in some specimens. Principal caudal-fin rays i,9,8,i* (36); caudal fin forked, lobes rounded, upper lobe slightly longer than lower lobe. Supraneurals 5 (1). Vertebrae 37 (1), with ribs present on vertebrae 5 to 24 (1). First dorsal-fin pterygiophore inserted posterior to neural spine of vertebra 10 (1). First analfin pterygiophore inserted posterior to haemal spine of vertebra 25 (1).
Color in alcohol. Ground color of head and body beige or light brown and countershaded ( Fig. 5 View FIGURE 5 ). Posterior tip of upper lip conspicuously dark. Snout with two dark brown stripes, one from nape to eye passing dorsally to posterior nostril, one from posterior tip of upper lip to eye; stripes present in specimens of all sizes, although inconspicuous in larger specimens. Upper half of opercle with two dark blotches, one at middle portion, the other at upper portion; blotches present in specimens of all sizes, although inconspicuous in some specimens. Dorsal portion of head with one dark brown transversal band at interorbital region and another on supraoccipital in all examined specimens irrespective of size, although inconspicuous in some specimens. Body scales with dark brown chromatophores concentrated on distal margin, resulting in overall reticulated pattern in specimens of all sizes; and with concentration of chromatophores on upper and lower portion of each body scale forming inconspicuous longitudinal lines in few specimens. In specimens of approximately 50 mm SL, body with eight to eleven dark brown transversal bars on dorsum, and several rounded blotches on flank, including one dark blotch on midline of anterior portion of the flank (between opercle and pelvic-fin origin) surrounded by five to seven dark blotches, one large blotch below dorsal fin, another above anal-fin origin, and a smaller at caudal peduncle. In specimens of 60 mm SL or more, body with ten to fourteen dark transversal bands on dorsum, and many rounded dark blotches on flank, including one dark blotch on midline of anterior portion of the flank (between opercle and pelvic-fin origin) surrounded by five to seven dark blotches, and three large midlateral ones below dorsal fin, above anal-fin origin and at caudal peduncle. Blotches on body varying in number from fifteen to approximately thirty; blotches also varying in shape from slightly elongate to rounded. Blotches formed exclusively by dermal pigmentation, except for three large midlateral blotches formed both by dermal and epidermal pigmentation. Dorsal, pectoral, pelvic, anal and caudal fins completely hyaline or with darkened rays in some specimens of 60 mm SL or larger. Caudal fin usually with uppermost and lowermost principal rays darkened. Adipose fin with base and distal margin darkened.
Color in life. Coloration in life similar to coloration in alcohol, except for head and body generally more countershaded and silvery; pectoral and pelvic fins yellowish to orange; anteriormost rays of dorsal, anal and caudal fins distinctly orange in some specimens; adipose fin reddish in some specimens, exposed posterior portion of body scales on dorsoanterior portion of flank in some specimens or of entire flank in others silvery ( Fig. 6 View FIGURE 6 ). Head and body completely dusky olive with clear spots on center of each flank scales in some large specimens.
Geographical distribution. Leporinus torrenticola is known only from the rapids of the middle rio Xingu and its main tributaries, the rio Iriri and rio Curuá, Pará state, Brazil ( Fig. 4 View FIGURE 4 ).
Etymology. From the Latin torrens meaning rushing waters, and incola meaning inhabitant, in reference of the preference of Leporinus torrenticola for fast flowing waters. An adjective.
Habitat and ecological notes. Our observations in the field corroborate Zuanon (1999), who states that Leporinus torrenticola preferably inhabits the moderate to fast flowing waters of the rapids of the rio Xingu and rio Iriri, where it is usually collected alongside specimens of Hypomasticus julii . Zuanon (1999) provided a beautiful study on the behavior and diet of this species, considering it omnivorous, feeding mainly on aquatic insects, filamentous algae and Podostemaceae .
Conservation status. Leporinus torrenticola is apparently endemic to the rapids of the rio Xingu and its main tributaries, occupying a relatively restricted area at northern Brazil. Part of the rapids of the rio Xingu was recently modified by the construction of Belo Monte, one of the largest hydroelectric dams in the world. The impacts of the Belo Monte to the populations of Leporinus torrenticola still need to be determined, even though it is expected to be severe due to species preference for rapids. Similarly, the distribution of this species is not quite well understood, and the species may occur in other portions of the rio Xingu and rio Iriri further away from the city of Altamira, where only few collections were made. More data on the distribution of the species and on how the Belo Monte dam is impacting the populations of the species are necessary to estimate the current conservation status of L. torrenticola . Therefore, according to the International Union for Conservation of Nature (IUCN) categories and criteria (IUCN Standards and Petitions Subcommittee, 2014), Leporinus torrenticola is better classified as Data Deficient (DD).
Remarks. The examined specimens of Leporinus torrenticola show great variation in the shape of the dorsal, pelvic, and anal fin, as well as in the thickness of the pectoral, pelvic and caudal fin rays and coloration of head, body and fins. Some specimens have dorsal, pelvic, and anal fins with distal margin distinctly concave (instead of rounded), pectoral, pelvic and caudal fins thickened, and head, body and fins generally darkened ( Fig. 8 View FIGURE 8 ). In those specimens, the anal fin is distinctly concave, with anterior branched rays approximately three times longer than last branched anal-fin ray. The dark brown blotches on these specimens are also arranged slightly different with the center of the midline blotch of the anterior portion of flank slightly ventral to the lateral line (instead of centered on the lateral line). However, these variations apparently occur irrespective of size, as there are specimens of similar sizes collected sintopically (e.g., INPA 31011, 87.5 and 92.7 mm SL) that show fins shaped differently, bearing fin rays of distinct thickness, and body colored distinctly. These variations apparently are not related to sexual dimorphism, since there are both mature males (MZUSP 96967, 115.8 mm SL) and females (MZUSP 96967, 125.0 mm SL) with convex fins. The significance of this variation is yet to be determined.
Due to these differences in the examined material of L. torrenticola , the type specimens were all selected among those collected in the rio Curuá, a tributary of the rio Iriri, itself a tributary of the rio Xingu, a geographical subset of the entire distribution of the species. All specimens collected in the rio Curuá are similar to each other ( Figs. 5 View FIGURE 5 , 6 View FIGURE 6 ) and none have the aforementioned variations in anal fin shape and coloration ( Fig. 8 View FIGURE 8 ).
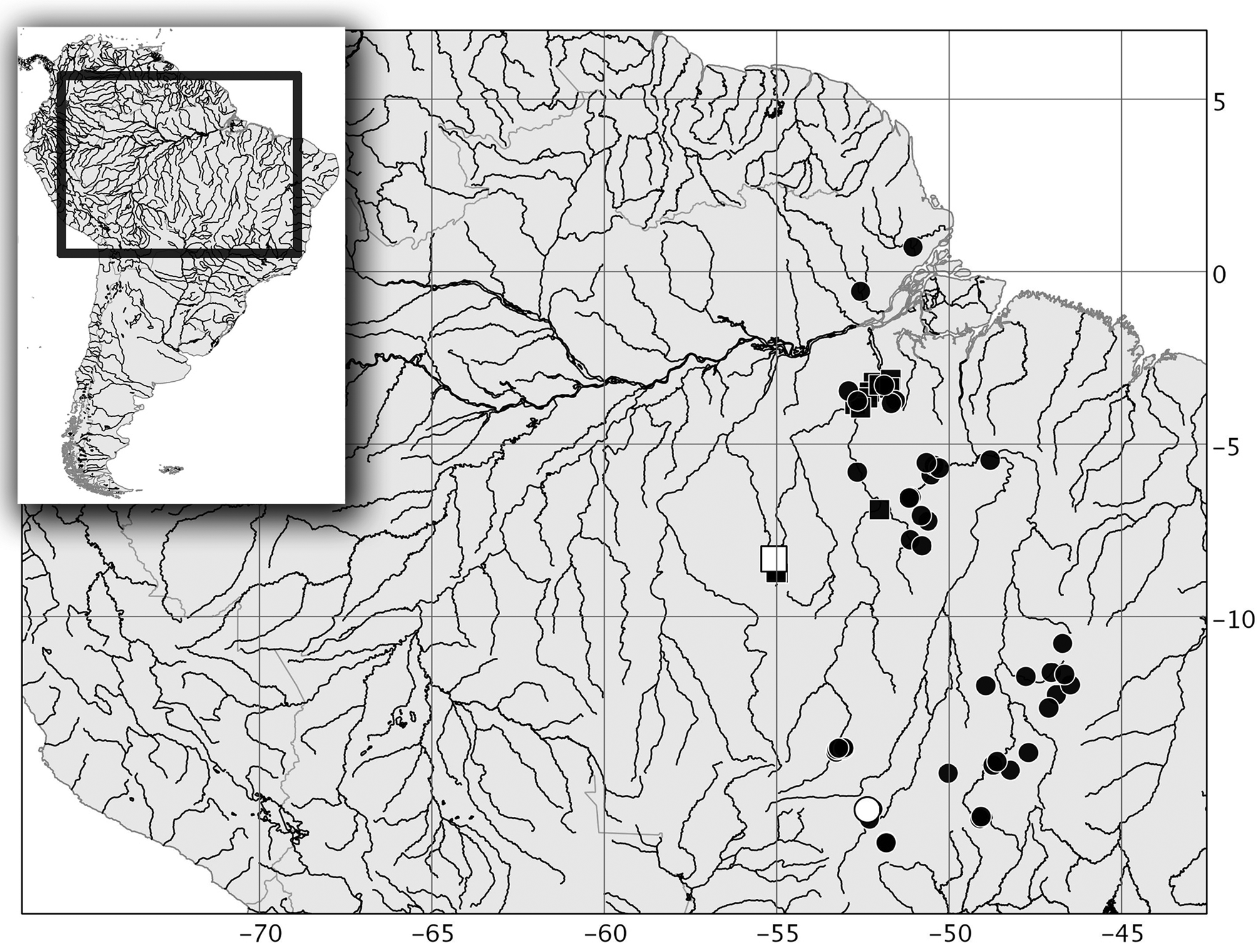




TABLE 2. Morphometric data for Leporinus torrenticola. SD = standard deviation.
| n | mean | range | SD | holotype | ||
|---|---|---|---|---|---|---|
| Standard length (SL) | 30 | 95.3 | 47.9 | 123.2 | 92.9 | |
| Percents of SL | ||||||
| Body depth | 30 | 25.1 | 21.4 | 28.0 | 1.7 | 25.0 |
| Caudal peduncle length | 30 | 9.4 | 8.5 | 10.4 | 0.5 | 10.2 |
| Head length | 30 | 23.2 | 21.5 | 24.8 | 0.8 | 22.6 |
| Predorsal distance | 30 | 47.2 | 45.4 | 49.1 | 1.0 | 47.4 |
| Prepelvic distance | 30 | 49.9 | 47.9 | 53.8 | 1.4 | 48.3 |
| Percents of head length | ||||||
| Eye diameter | 30 | 27 | 23 | 31 | 2.1 | 25 |
| Snout length | 30 | 40 | 35 | 45 | 2.3 | 41 |
| Bony interorbital | 30 | 38 | 33 | 42 | 1.9 | 42 |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Leporinus torrenticola
| Birindelli, José L. O., Teixeira, Tulio F. & Britski, Heraldo A. 2016 |
Leporinus
| Zuanon 1999: 87 |