
Choniognathus Rathbun, 1932
|
publication ID |
https://doi.org/ 10.11646/zootaxa.5048.3.1 |
|
publication LSID |
lsid:zoobank.org:pub:951BE302-C0BF-4AA3-AE12-BBAC4EDEBAFB |
|
DOI |
https://doi.org/10.5281/zenodo.5569443 |
|
persistent identifier |
https://treatment.plazi.org/id/03B187B7-FF95-FFB9-89B3-A1D5FAC65FC8 |
|
treatment provided by |
Plazi |
|
scientific name |
Choniognathus Rathbun, 1932 |
| status |
|
Genus Choniognathus Rathbun, 1932 View in CoL
Choniognathus Rathbun, 1932: 33 View in CoL .— Sakai, 1938: 267 (key), 272, 273.— Griffin & Tranter, 1986: 200 (key), 201–203.
Material examined. Holotype of Choniognathus koreensis Rathbun, 1932 : female (cl 9.1 mm, pcl 8.3 mm, cw 6.0 mm, bcw 5.3 mm) ( USNM 48204 About USNM ), stn 4879, Oki Shima , Korea, 3417’ N 13015 View Materials ’E, 108 m, fine gray sand, broken shells, coll. Albatross, 2 August 1906 . Other material of Choniognathus koreensis Rathbun, 1932: 1 female (poor condition) (cl 9.1 mm, pcl 8.3 mm, cw 6.2 mm, bcw 5.3 mm) ( KPM NH 0104599 , part), Manazuru Town, Ashigarashimo District, Sagami Bay , Japan, coll. T. Sakai , 1962.—1 ovigerous female (cl 6.3 mm, pcl 6.0 mm, cw 4.0 mm, bcw 3.6 mm) ( ZRC 2020.373 View Materials ), station T36 , Cortes , Panglao , Bohol, Visayas , 123–135 m, 943.3’N 12348.8’E, Philippines, on mud, coll. PANGLAO 2004, 24 June 2004.
Type species. Choniognathus koreensis Rathbun, 1932 View in CoL , by monotypy
Diagnosis. Carapace and pereopod surfaces usually covered by dense short tomentum which completely obscures margins. Carapace longitudinally ovate, constricted behind hepatic regions. Regions clearly demarcated, raised; surface covered by many small tubercles, granules and/or short spines. Pseudorostrum short, usually with wide U-shaped cleft. Supraorbital eave wide curved, preorbital spine formed by proximal edge of eave; 1 small intercalated triangular spine, usually separated from adjacent structures by visible gap; postorbital and suborbital lobes surround orbit. Hepatic region with tubercles and granules, sometimes spines; gastric region raised with large granules or tubercles; cardiac region prominently raised with 1 large median tubercle; branchial region raised, covered with granules and tubercles; metabranchial region with 1 blunt tubercle. Antennal flagellum very short, not extending beyond short pseudorostral horns. Basal antennal article broad short covered by short setae. Third maxilliped with merus and ischium smooth or covered with tubercles, merus may be fused with ischium; merus with anteroexternal angle produced to differing degrees. Cheliped short; dorsal margins of carpus, propodus and chela unarmed or with spines. Ambulatory legs short without spines; dactylus relatively long, sharp. Male and female pleons with 6 free somites and telson; narrow in male; somite 1 with median swelling.
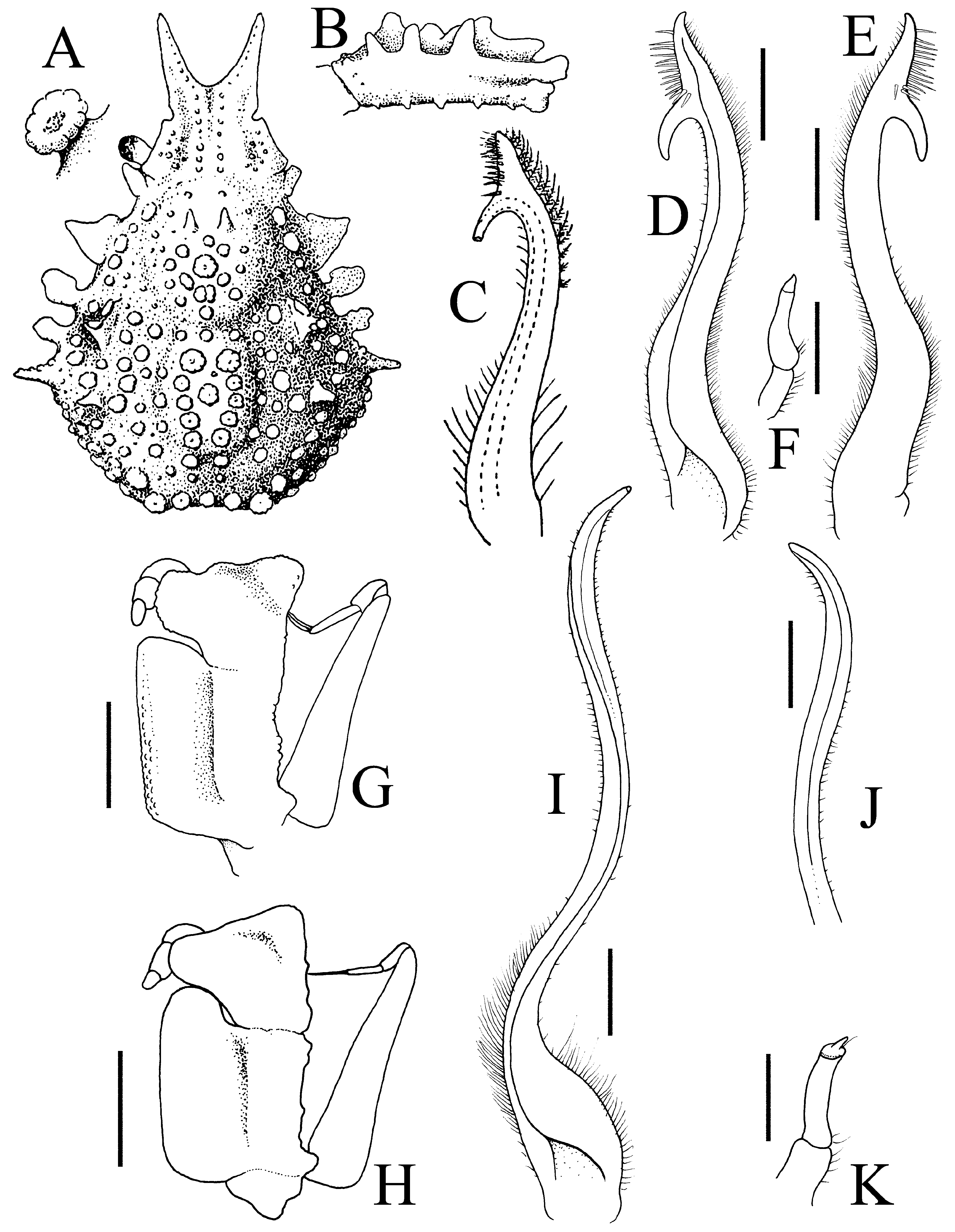
Remarks. Rathbun (1932) established Choniognathus on the basis of a new species, C. koreensis Rathbun, 1932 , described from one female from the Sea of Japan. Sakai (1938: 273) argued that this species is probably identical with Eurynome reini Balss, 1924 , described from two females from Sagami Bay. The two species are now regarded as synonyms (see Yokoya 1933: 158; Sakai 1965: 78; Takeda & Miyake 1969: 511; Sakai 1976: 222; Ng et al. 2008: 116). The holotype female of C. koreensis ( Figs. 7A, B View FIGURE 7 , 13G View FIGURE 13 ) was examined and Sakai’s (1938) decision is supported. The only major difference is that the anterointernal angle of the ischium of the holotype of C. koreensis is somewhat lower and less auriculiform in shape ( Fig. 13G View FIGURE 13 ) whereas it is more developed in other specimens ( Fig. 13H View FIGURE 13 ).
A major diagnostic character for Choniognathus is that the merus and ischium of the third maxillipeds are fused, first figured by Sakai (1938: text-fig. 34b) (see also Sakai 1976: text-fig. 120b) ( Fig. 13G, H View FIGURE 13 ). The third maxillipeds of the present specimens of C. reini confirm this ( Fig. 13G, H View FIGURE 13 ). Griffin (1965) re-examined the holotype female and other specimens of C. granulosus and described and figured the species at length. He noted that it is different from C. reini in that the pseudorostral spines are more divergent ( Griffin 1965: fig. 1), the hepatic and branchial regions are proportionately wider ( Griffin 1965: fig. 1), and the merus and ischium of the third maxilliped are separated by a distinct suture and likely to be mobile ( Griffin 1965: fig. 2). Griffin (1965: 36), however, did comment that “Therefore it is important to note that in E. granulosa , while these two segments of the maxillipeds are distinct, the junction seems a very narrow, shallow, groove and it is possible that the merus is not freely moveable”. Griffin & Tranter (1986) later stated that the merus and ischium of the third maxilliped is not fused in E. granulosus , and that this is not a diagnostic character for Choniognathus . Griffin & Tranter (1986) also transferred Eurynome elegans and E. verhoeffi to Choniognathus , but without much explanation.
The condition of the third maxilliped merus and ischium in C. granulosus should be re-examined to ascertain if it is actually mobile, especially as Griffin (1965) had noted the suture was shallow. It is known that for pleonal somites in some Xanthidae and Geryonidae , the presence of a suture is not always associated with mobility, and the structures may be functionally fused (e.g., see Ng & Chia 1994; Ng et al. 1998; Manuel-Santos & Ng 2007). In C. spinosus n. sp., the merus and ischium of the third maxillipeds are clearly mobile ( Fig. 6D View FIGURE 6 ).
The G1 of C. reini has never been figured but it is not of the typical Eurynome type; Griffin & Tranter (1986: 203) describing it as broad, straight and distally it is curved strongly ventrally and slightly outwards; apex blunt with long setae on the medial margin and a blunt subterminal lobe on the lateral margin; aperture is a narrow slit-like opening on the sternal surface of this subterminal lobe..
Choniognathus verhoeffi described from two males and two females from the Red Sea ( Balss 1929: 12, fig. 5) does not belong to the genus. The frontal margin is quadrilobate, with a pair of median spines and strong, anteriorly directed lateral spines, the dorsal surface of the carapace is covered by regularly spaced boletiform tubercles, the antennal flagellum is very long, being much longer than the pseudorostral spines, the chelipeds are elongate and the outer surfaces are strongly spinate, the meri and carpi of the ambulatory legs are long, prominently spinate, and the male telson is linguiform and long ( Balss 1929: fig. 5). Griffin & Tranter (1974: 168, fig. 1d, e) discussed C. verhoeffi with fresh material (one male and three females from the Gulf of Eilat, Red Sea, 68–90 m) and described the G1 as follows: “first pleopod of the male is expanded distally but the opening is simple (but with no figure). They noted that it was quite different from the “scyriform G 1 type of typical Eurynome species ( Griffin & Tranter 1974: 168). As far as is known, C. verhoeffi is also not covered with dense tomentum which completely obscures the surfaces and outlines like in C. reini . It was almost certainly on this character that they decided to later transfer it to Choniognathus ( Griffin & Tranter, 1986: 203) . The species certainly cannot be assigned to any of the genera discussed in this paper. It is provisionally retained in Choniognathus until the types can be re-examined.


| KPM |
Kanagawa Prefectural Museum of Natural History |
| T |
Tavera, Department of Geology and Geophysics |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
InfraOrder |
Brachyura |
|
SuperFamily |
Majoidea |
|
Family |
Choniognathus Rathbun, 1932
| Forges, Bertrand Richer De, Lee, Bee Yan & Ng, Peter K. L. 2021 |
Choniognathus
| Griffin, D. J. G. & Tranter, H. A. 1986: 200 |
| Sakai, T. 1938: 267 |
| Rathbun, M. J. 1932: 33 |