
Vansoestia, Díaz, Maria C., Thacker, Robert W., Redmond, Niamh E., Perez, Thierry & Collins, Allen G., 2015
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3956.3.5 |
|
publication LSID |
lsid:zoobank.org:pub:25F9FA82-28BA-4C94-8EF5-D0EE88B0B48E |
|
DOI |
https://doi.org/10.5281/zenodo.6093271 |
|
persistent identifier |
https://treatment.plazi.org/id/03AF87AD-1138-6354-84A1-FC4EFA80529F |
|
treatment provided by |
Plazi |
|
scientific name |
Vansoestia |
| status |
gen. nov. |
Vansoestia View in CoL , gen. nov.
Diagnosis. Ianthellidae represented by individuals with a thin (1–4 mm) encrusting growth form that presents a cellularized cortex of variable thickness (10–200 µm) composed of a collagenous matrix and various types of cells with inclusions (i.e. granular, spherulous, and/or vacuolar). Abundant subdermal canals and lacunae are found immediately underneath the cortex. The choanosome consists of numerous aquiferous canals, a mesohyl where two or three types of cells with inclusions can be distinguished, and oval eurypylous choanocyte chambers 20–50 µm, all supported with abundant fibrillar collagen.
Type species: Vansoestia caribensis gen. nov. sp. nov.
Material examined. All specimens studied were found in coral reefs from Bocas del Toro, Panama, between 10–20 m in depth. This species grows over dead coral and other organisms, in open and cryptic reef habitats. Holotype: USNM 1133773, Buoy 19 (N9°18.11, W 82°17.66), Bocas del Toro, Panama (P67), 10 m deep, coll: M.C. Díaz and R.W. Thacker; 7/18/2009
Paratypes: USNM 1133782, Buoy 19 (N9°18.11, W 82°17.66), Bocas del Toro, Panama (P76), 15 m deep, coll: M.C. Díaz and R.W. Thacker; 7/18/2009; USNM 1204851 (P 12x 342), Punta Caracol (N9°22.66, W 82°18.19), 15 m deep, coll: M.C. Díaz and S.A. Pomponi, 8/8/2012.
Additional material. P 12x 402, and P 12x 403 were consumed in histological, or TEM preparations.
Etymology. The species is named in honor of Dr. Rob van Soest, an essential contributor to the understanding of Demospongiae, including Verongida , in the Caribbean and worldwide.
Description. External morphology: The sponge is soft, thinly encrusting (1–4 mm thick), orange to yellow in color internally and externally. Color turns slowly brown to purple in ethanol. The sponge grows over dead coral or over other organisms, particularly other sponges ( Figure 1 View FIGURE 1 ). Its surface is dominated by thin reticulated subdermal canals (3–8 mm wide) that branch and decrease in width away from the oscules (0.5–1 cm wide). Small ostia (0.4– 0.8 mm wide) are dispersed between subdermal canals. The soft and fragile consistency of this sponge reflects the lack of a reticulate fiber skeleton.
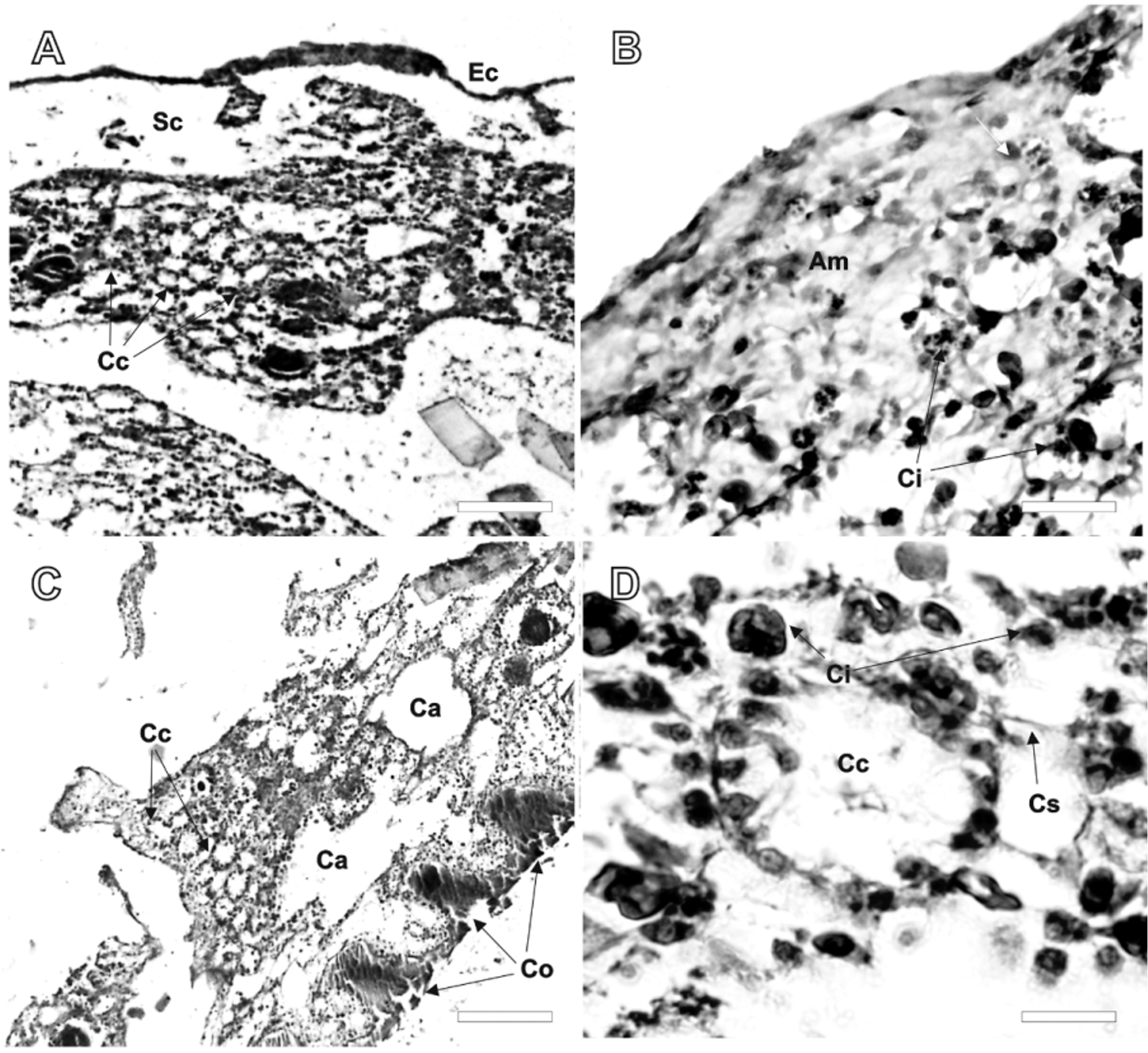
Internal morphology ( Figures 2–4 View FIGURE 2 View FIGURE 3 View FIGURE 4 ). There is a clear separation between ectosome and choanosome ( Figure 2 View FIGURE 2 A). The ectosome is a collagenous cortex of variable thickness (10–200 µm), with collagen in the form of an amorphous matrix containing loose cells with inclusions ( Figure 2 View FIGURE 2 A, B), and an external cuticle ( Figure 3 View FIGURE 3 A). Large channels and lacunae (subdermal spaces) are observed immediately underneath the ectosome ( Figure 2 View FIGURE 2 A). The choanosome consists of densely arranged oval-shaped choanocyte chambers (20–40 µm in diameter) ( Figure 2 View FIGURE 2 A, C), and a mesohyl reinforced by collagen fibrils arranged in strands ( Figure 3 View FIGURE 3 B, C), packed with various types of cells with inclusions, and with abundant aquiferous canals. The chambers are densely and homogeneously distributed in some areas ( Figure 2 View FIGURE 2 A, C), and rare in others that are instead densely packed by conglomerates of cells ( Figure 3 View FIGURE 3 D). Choanocytes (4–6 µm in diameter) appear oval to triangular in shape ( Figure 2 View FIGURE 2 D). Larger cells with inclusions of various shapes and sizes (4–12 µm in diameter) can be observed in both the mesohyl and the ectosome ( Figures 2–3 View FIGURE 2 View FIGURE 3 ). The largest cells with inclusions (Ci1) are widespread in the mesohyl, in the cortex, and lining canals ( Figure 3 View FIGURE 3 B and 3D). There are some canals lined by these large cells with inclusions ( Figure 3 View FIGURE 3 D), and others without them ( Figure 3 View FIGURE 3 C).
TEM sections allowed further distinction of the various types of cells with inclusions, and the observation of archaeocytes, choanocytes, and collagen fibrils. Spherulous cells 1 ( Figure 4 View FIGURE 4 A, B and D) are ovoid to roundish slightly deformed (6–10 µm in diameter) with spherules (0.7–2 µm wide), containing microgranular inclusions that are dense but grey or black in color (0.06–0.15 µm in diameter), with a clear round nucleus (1.3–2 µm in diameter). By the size and distribution of spherules, these cells probably correspond to the cells with inclusions 1 (Sc1) observed in the histological cross sections ( Figures 2–3 View FIGURE 2 View FIGURE 3 ). Spherulous cells 2 ( Figure 4 View FIGURE 4 B) are similar in size and shape of Sc1, but their spherules are larger and ellipsoid in shape (0.1–0.3 µm in diameter), and electron dense dark. These cells might represent a different stage of the spherulous cell 1. A third type, which seems like vacuolar cells ( Figure 4 View FIGURE 4 C), occurs with variable shapes, smaller size (5–8 µm in diameter), and containing a single round, dark dense granule (0.4–0.8 µm in diameter) within each vacuole (1–1.5 µm in diameter). By the shape and size of the granules, these cells likely correspond to the cell with inclusion 2 observed in the histological cross sections. Finally, a microgranular cell with membrane-bound inclusions with striated content, was observed once ( Figure 4 View FIGURE 4 D). A thick layer of collagen fibrils (Co) with loose bacteria surrounds the choanocyte chambers and long cytoplasmic projections could be seen in some choanocytes ( Figure 4 View FIGURE 4 E). One choanocyte with a basal nucleolated nucleus was observed ( Figure 4 View FIGURE 4 F).
Remarks. The delicate and thin specimens of Vansoestia caribensis have been found in Bocas del Toro, Panama, since 2006; however, only the slow oxidation observed when specimens were placed into alcohol suggested an affinity to verongids. Due to the lack of a fiber skeleton, we initially affiliated this sponge to the genus Hexadella . Histological observations corroborated its ianthellid nature (large, sac-shaped choanocyte chambers), but its distinct identity and closest affinity to the known sequences of fiber-bearing ianthellid genera from the Western Tropical Pacific appeared after analyzing 18S and 28S ribosomal gene sequences (Redmond et al., 2013; Thacker et al., 2013). Its genetic distance from the only previously known fiber-less verongid genus, Hexadella , prompted the need to erect a new genus to classify this species. This biological record constitutes the first member of family Ianthellidae ever encountered in the Caribbean Sea.
There is a striking similarity between V. caribensis gen nov. sp. nov. and the external appearance of Hexadella pruvoti Topsent, 1896 ( Reveillaud et al., 2012) , particularly with respect to the abundant subdermal canals converging in a wide oscule, and its yellowish coloration. However, 18S and 28S gene sequence analyses confirmed the very distinct nature of these two species. A cellularized cortex of variable thickness (10–200 µm thick) composed of amorphous collagen and various types of cells is present in V. cari bensis gen. nov. sp. nov. and forms an important morphological difference between these two genera, since Hexadella possesses a thinner cortex with a 1 µm thick cuticle. A well-developed cellularized cortex might represent a potential synapomorphy between Vansoestia gen. nov., Ianthella , and Anomoianthella ( Bergquist, 1980, 1995; Bergquist and Kelly-Borges, 1995; Bergquist and Cook, 2002; present study). Indeed, Bergquist, (1995) and Bergquist and Kelly-Borges (1995) described a collagenous ectosome (70 to 300 µm thick) containing cells for all Ianthella species and a cellularized cuticle for Anomoianthella species ( Bergquist, 1980). Together with the presence of a collagen-reinforced cortex combined with a cuticle, the high degree of reinforcement of the choanosome with collagen is also a characteristic shared by Vansoestia gen. nov., Ianthella and Anomoianthella .
Some similarities among the spherulous cells of V. caribensis gen. nov. sp. nov., and other verongids were also noticed ( Vacelet, 1967). The cells with striated membrane-bound bodies are very similar to cells observed in Aplysina aerophoba by Maldonado (2009). The dark granules found in cells of Vansoestia gen. nov. might represent glycogen granules ( Maldonado, 2009), or they might contain brominated compounds (Thompson, 1986) like has been found for microgranular and spherulous cells of two different Aplysina species. The large diversity of cells with inclusions, and their similarities with species of Hexadella and other verongid genera invites continued research to understand the morphology and physiological functions of these cells and their products.
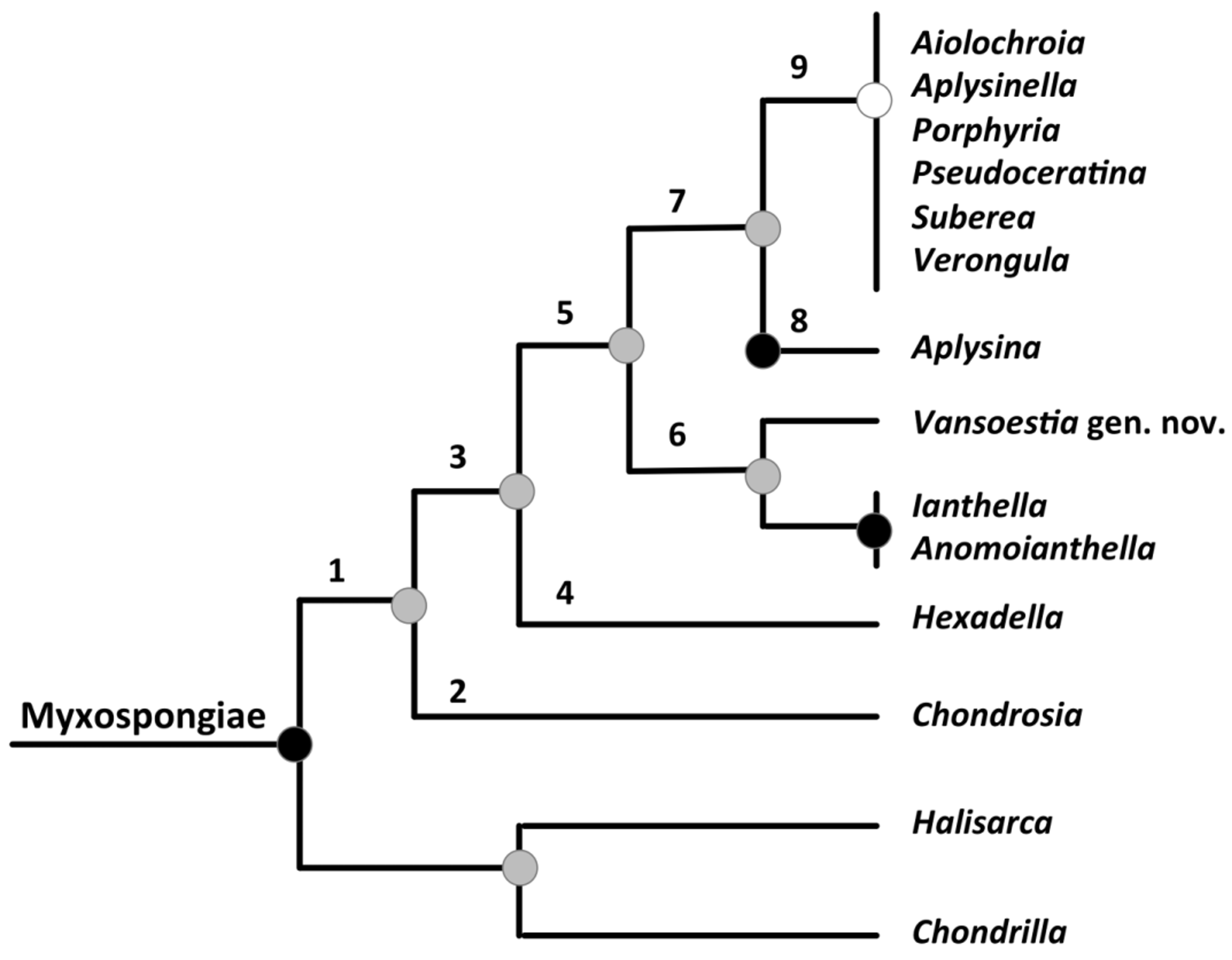
Phylogenetic remarks. 18S and 28S sequences demonstrate that this fiber-less member of Ianthellidae is a sister group to the clade that includes Ianthella and Anomoianthella . (Redmond et al., 2013; Thacker et al., 2013). Figure 5 View FIGURE 5 summarizes the current phylogenetic interpretation of the relationships among verongid genera based on the latest molecular, and morphologic studies of the group ( Erwin and Thacker, 2007; Erpenbeck et al., 2012; Díaz et al., 2013; Redmond et al., 2013; Thacker et al., 2013). The two sister clades make up the subclass Myxospongiae, one containing the amended Verongida (after Redmond et al., 2013) and the other containing Chondrilla and Halisarca (Order Chondrillida sensu Redmond et al., 2013). All verongids are grouped in a strongly supported monophyletic clade ( Figure 5 View FIGURE 5 , clade 1), which shares the presence of collagen fibrils forming bundles either in the cortex, in the mesohyl, or within fibers. Chondrosia (family Chondrosiidae ) forms a wellsupported sister clade to all other verongids ( Figure 5 View FIGURE 5 , clade 2), and is represented by sponges with extremely high collagen content, a thick (> 1 mm thick) “collagenous cortex”, and a fiber-less body. The rest of the verongids form a highly supported clade ( Figure 5 View FIGURE 5 , clade 3), containing all taxa that produce brominated secondary metabolites and have abundant spherulous cells. Basal within this group, a highly supported clade ( Figure 5 View FIGURE 5 , clade 4) contains all Hexadella species which share eurypylous choanocyte chambers, a thin cuticle (1–2 µm) and a very thin ectosome. The next clade ( Figure 5 View FIGURE 5 , clade 5) is a moderately supported clade containing all non- Hexadella verongids. Within clade 5, two clades are distinguished: a moderately (85%) supported clade ( Figure 5 View FIGURE 5 , clade 6) containing Vansoestia gen. nov. (skeleton less) and the fiber bearing Pacific taxa of genera Ianthella and Anamoianthella, and a moderately to highly supported clade ( Figure 5 View FIGURE 5 , clade 7) including all the taxa possessing diplodal choanocyte chambers. Within clade 7 ( Figure 5 View FIGURE 5 ), a clearly monophyletic group of sponges (clade 8) contains all Aplysina species, whereas a poorly supported clade (clade 9) contains at least six genera ( Aiolochroia , Aplysinella , Porphyria , Pseudoceratina , Suberea , Verongula ) and an undescribed genus from Moorea (Díaz et al., 2013).
The Verongida taxa with eurypylous, sac-shaped choanocyte chambers are placed in two distinct clades ( Figure 5 View FIGURE 5 , clades 4 and 6), demonstrating that this feature is paraphyletic within Verongida . Current studies suggest the classification of Verongida comprises four main clades: (1) a clear monophyletic group containing all Aplysina species studied, and representatives of all Aplysinellidae , and Pseudoceratiniidae genera; (2) a clade that contains Ianthella , Anomoianthella , and Vansoestia gen. nov., but excludes Hexadella , which should be defined by an amended family Ianthellidae ; (3) a clade that presently contains only Hexadella and that should be erected as a new family; and (4) a clade containing the family Chondrosiidae sensu Redmond et al. (2013) .
We demonstrate here that the suprageneric classification of Verongida must be further revised in order to reflect our current knowledge of phylogenetic relationships among the genera of Myxospongiae. Considering the diagnostic characters that we have in hand, a thorough comparative review of the histological and ultrastructural features of representatives of each genus is definitely needed to improve the taxonomic diagnoses of these taxa.



| USNM |
Smithsonian Institution, National Museum of Natural History |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |