
Oxynoemacheilus phasicus, Freyhof & Kaya & Epitashvili & Geiger, 2021
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4952.1.8 |
|
publication LSID |
lsid:zoobank.org:pub:6EC786D4-DF85-419A-B350-8CCC2C487CD1 |
|
DOI |
https://doi.org/10.5281/zenodo.4685572 |
|
persistent identifier |
https://treatment.plazi.org/id/03AEEE34-FFB8-C800-FF0A-FF75FE37F895 |
|
treatment provided by |
Plazi |
|
scientific name |
Oxynoemacheilus phasicus |
| status |
sp. nov. |
Oxynoemacheilus phasicus , new species
( Figs 2–5 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 )
Holotype. ZFMK ICH-124180 , 74 mm SL; Georgia: Dzirula River at road from Tbilisi to Kutaisi 1 km upstream from Khunevi, 42.0975 43.3900. GoogleMaps
Paratypes. FSJF 3592 , 1 , 77 mm SL; same data as holotype GoogleMaps .— FSJF 3576 , 3 , 50–59 mm SL; Georgia: Enguri River at Lia, 42.6617 41.9911 GoogleMaps .— FSJF 3615 , 2 , 44–54 mm SL; Georgia: Tskhenitskali River upstream of Samtredia, 42.1694 42.4314 GoogleMaps .— FSJF 4061 , 2 , 32–39 mm SL; Georgia: Gubistskali River east of Samtredia , upriver of confluence with Rioni and Rioni channel, 42.16251 42.41069 GoogleMaps .
Material used in molecular genetic analysis. ZFMK ICH-NB184 ; same data as holotype ( GenBank accession number: MW 564259 View Materials , holotype or paratype, association unclear) GoogleMaps .— ZFMK ICH-106137-106138 ; Georgia: Swlana stream west of Jurukveti, 42.07829 41.91643 (GenBank accession numbers: MW564307 View Materials , MW564435 View Materials ) GoogleMaps .— ZFMK ICH-106139-106140 ; Georgia: Gubistskali River east of Samtredia , upriver of confluence with Rioni and Rioni channel, 42.16251 42.41069. (GenBank accession numbers: MW564371 View Materials , MW564381 View Materials ) GoogleMaps .— ZFMK ICH-NB190 - NB191; Georgia: Tskhenitskali River upstream of Samtredia, 42.1694 42.4314 (GenBank accession numbers: MW564373 View Materials , MW564302 View Materials , MW564481 View Materials ) GoogleMaps .— ZFMK ICH-NB195 ; Georgia: Enguri River at Lia, 42.6617 41.9911 (GenBank accession number: MW 564442 View Materials ) GoogleMaps .
Diagnosis. Oxynoemacheilus phasicus is distinguished from O. cemali , the only other species of Oxynoemacheilus known from the eastern Black Sea basin, by a slightly emarginate caudal fin (vs. deeply emarginate), pelvic fin not reaching to origin of genital papillae (vs. reaching to or beyond posterior tip of genital papillae), pelvic-fin origin below first branched dorsal-fin ray (vs. below second or third ray), a wide band in posterior half of caudal fin (vs. absent), and a mottled or marbled flank pattern (juveniles with bars on flank behind dorsal-fin base) (vs. usually with 9–15 narrow, irregularly shaped and spaced, dark-grey bars or vertically elongate blotches on flank posterior to dorsal-fin origin).
Oxynoemacheilus phasicus is distinguished from the other members of the O. brandtii species group ( O. brandtii , O. elsae , and O. merga ) by having a slightly emarginate caudal fin (vs. forked in O. brandtii , deeply emarginate in O. elsae and O. merga ), pelvic fin not reaching to origin of genital papillae (vs. reaching to or beyond anus in O. brandtii and many O. elsae and O. merga ), caudal peduncle 1.5–2.1 times longer than deep (vs. 2.5–3.2 in O. brandtii , 2.3–2.9 in O. elsae , 2.0– 2.5 in O. merga ), and a mottled or marbled flank pattern (vs. usually with irregularly shaped and spaced, dark-brown bars or vertically elongate blotches on flank posterior to dorsal-fin origin, much wider than interspaces, often fused into a plain brown pattern on flank in front of dorsal-fin base in O. brandtii ; usually with irregularly shaped and spaced, rarely regularly shaped and spaced, dark-brown bars, often split in middle, or flank almost plain brown in O. elsae ; with many, narrow, pale-brown bars, usually much more narrow than interspaces, often dissociated in vertically elongate blotches on flank and dorsal saddles, rarely mottled in O. merga ).
Description. See Figures 2–5 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 for general appearance and Table 2 View TABLE 2 for morphometric data. Medium-sized and moderately slender species. Body deepest at about midline between nape and dorsal-fin origin. Body width greatest at pectoral-fin base, at cheek in largest individual. Section of head roundish, flattened on ventral surface, slightly convex in interorbital space, convex on snout. Snout blunt. Caudal peduncle compressed laterally, 1.5–2.1 times longer than deep. Pelvic axillary lobe present, its tip attached to flank. Pelvic-fin origin below first branched dorsalfin ray. Anal-fin origin at about vertical of midline between dorsal and caudal-fin origins. Pectoral fin reaching to approximately 50–70% of distance from pectoral-fin origin to pelvic-fin origin. Pelvic fin not reaching anus; reaching slightly anterior to vertical of tip of last dorsal-fin ray; almost reaching anus in juveniles smaller than 40 mm SL. Anus about 30–70% of an eye diameter anterior to anal-fin origin. Anal fin not reaching caudal-fin base. No dorsal or ventral adipose crest on caudal peduncle. Largest known individual 77 mm SL.
Dorsal fin with 8½–9½ branched rays, outer margin straight or convex. Anal fin with 5½ branched rays, outer margin straight or slightly concave. Pectoral fin with 9–11 branched rays, outer margin straight. Pelvic fin with 7 branched rays, outer margin straight or slightly convex. Caudal fin slightly emarginate with 8+8 branched rays, moderately emarginate in juveniles smaller than 40 mm SL. Flank, back and abdomen densely covered by scales. Lateral line complete, terminating at caudal-fin base. Anterior nostril opening at end of a low, ovoid and flap-like tube. Posterior tip of anterior nostril overlapping in small individuals, not overlapping posterior nostril when folded backwards in large individuals. One central pore, two in one individual, and one lateral pore on each side of supra- temporal head canal, 8–10 pores in anterior infraorbital canal, 3–4 pores in posterior infraorbital canal, 6–9 pores in supraorbital canal and 5–7 pores in mandibular canal. A short suborbital groove in male. Mouth small, arched. Lips thick without furrows, lower lip thicker than upper lip. A median interruption in lower lip. Upper lip with a small and short median incision in some individuals, absent in others. Processus dentiformis narrow, very small and rounded. Lower jaw rounded, without median notch. Barbels long; inner rostral barbel reaching to or beyond base of maxillary barbel, outer reaching beyond anterior margin of eye, to middle of eye in some individuals. Maxillary barbel reaching beyond vertical through middle of eye or to or beyond posterior margin of eye. Barbels very short in largest individuals.
Coloration. Body with yellowish or pale brown background and dark- or pale-brown pattern in live and preserved individuals. Preserved individuals with a wide inner-axial stripe, absent in life. Dorsal head and upper part of cheek brown, with a vermiculate pattern. Ventral surface of head yellowish without pattern. Flank densely mottled, without bars or blotches, marbled or with bars on flank behind dorsal-fin origin in individuals smaller than 50 mm SL, almost plain brown in some individuals. Flank pigmentation reaching down to horizontal of pectoral-fin base or to belly in some individuals. Back mottled, marbled or with 3–4 squarish or triangular predorsal blotches, confluent with flank pattern, one saddle at dorsal-fin origin, one or none in middle of dorsal-fin base and one at end of dorsal- fin base, and 3–6 saddles behind dorsal-fin base, all saddles wider than interspaces. A narrow, black bar at caudal-fin base. Posteriormost upper and lowermost caudal peduncle with a pale yellowish, unpigmented blotch. Dorsal and pectoral fins with small brown blotches on rays, forming 1–3 bands. Caudal fin with many small brown blotches on rays, forming 1–3 wide and bold bands, often band at second half of caudal fin very wide or caudal-fin rays almost completely brown. Pectoral, anal and pelvic fins with dark-brown blotches on rays; hyaline or with few blotches in individuals smaller than 60 mm SL.
Distribution. Oxynoemacheilus phasicus was collected from the Enguri and Rioni River drainages in Georgia. It has not been found in the Chorokhi / Çoruh River in Georgia or Turkey or in small rivers in-between despite intensive field work in that area.
Etymology. The species name is derived from Phasis , the Greek name of the Rioni. An adjective.
Remarks. Nemacheilus bergi was named by Gratzianov (1907) based on a description by Berg (1899) of two individuals (61 mm, 35 mm) from the Akstapha [Agstafa] River, a right tributary of the Kura in Azerbaijan. Berg (1899:73) wrote in his chapter on Nemacheilus sp.: „Die Schwanzflosse ist im Verhältnis zu N. Brandt schwach ausgeschnitten (bei dem Exemplar von 61 mm. Länge ist der mittelste Strahl der Schwanzflosse 8 mm. Und die seitlichen 11 mm lang).” [The caudal fin is slightly emarginate in comparison with N. brandtii (in the specimen of 61 mm the length of the central caudal-fin ray is 8 mm and of the lateral ones 11 mm)]. Also, Berg (1899:72–73) mentions that the pelvic fin reaches the anus, there is a bold bar at the caudal-fin base, a wide band in the middle of the caudal fin and incomplete bars on the flank. This species is further treated as a synonym of O. angorae by Berg (1949), who distinguished O. angorae from O. bergianus by the caudal peduncle being less than two times longer than deep in O. angorae vs. more in O. bergianus . Çiçek et al. (2018) treat N. bergi as a valid species in Oxynoemacheilus without giving further details. Soheil Eagderi (Tehran) sent JF a picture ( Fig. 7 View FIGURE 7 ) of one individual they identified as O. bergi collected in the Kura, very close to the inflow of the Agstafa. Measured from the picture, this individual has the caudal-peduncle depth two times in the caudal-peduncle length, the caudal fin is emarginate and the middle caudal-fin ray is 75% of the length of the longest principal ray in the lower caudal-fin lobe (72% in the description of Berg 1899, but unclear which lobe was measured). These two character states as well as its colour pattern distinguish this species clearly from O. brandtii (caudal peduncle 2.5–3.2 times longer than deep, caudal fin forked). Indeed the fish on Fig. 7 View FIGURE 7 agree with the characters given for N. bergi by Berg (1899) and we support the view of Çiçek et al. (2018) treating O. bergi as a valid species.
Oxynoemacheilus bergi is clearly distinguished from O. phasicus by possessing a deeply emarginate caudal fin (vs. slightly emarginate in O. phasicus ), and the pelvic fin reaching the anus (vs. not reaching). Potentially O. bergi is conspecific with O. bergianus , described from the Sefid River in Iran and known to occur in the adjacent Aras, a major tributary of the Kura. Future research should clarify if O. bergi is different from O. bergianus . If both are conspecific, O. bergianus would become a junior synonym of O. bergi .
We also follow Çiçek et al. (2018) in treating N. angorae alasanicus as a synonym of O. bergi and N. brandtii gibbusnazus as a synonym of O. brandtii . Çiçek et al. (2018) did not examine any materials of N. a. alasanicus or N. b. gibbusnazus, both described by Elanidze (1983) from the upper Alazani in the Georgian Kura drainage. G. Epitashvili and L. Mumladze (Ilia State University, Tbilisi, Georgia) visited the upper Alazani though yet only O. brandtii was collected. Çiçek et al. (2018) give no arguments for their conclusion and did not analyse the description of both species. However, we also have no arguments to reject the hypothesis of Çiçek et al. (2018), as the descriptions are not very conclusive, without figures and we also did not examine enough materials by our own. Only four loaches from the type locality of both species were available for this study and these are identified as O. brandtii . It is likely that Elanidze (1983) had identified his materials by the key provided by Berg (1949) and correctly identified one species as O. brandtii and the second species, which has a deeper caudal peduncle and an emarginate (vs. furcate) fin, was identified as O. angorae . He then described new subspecies for both species, and thus N. angorae alasanicus might be identified as O. bergi .
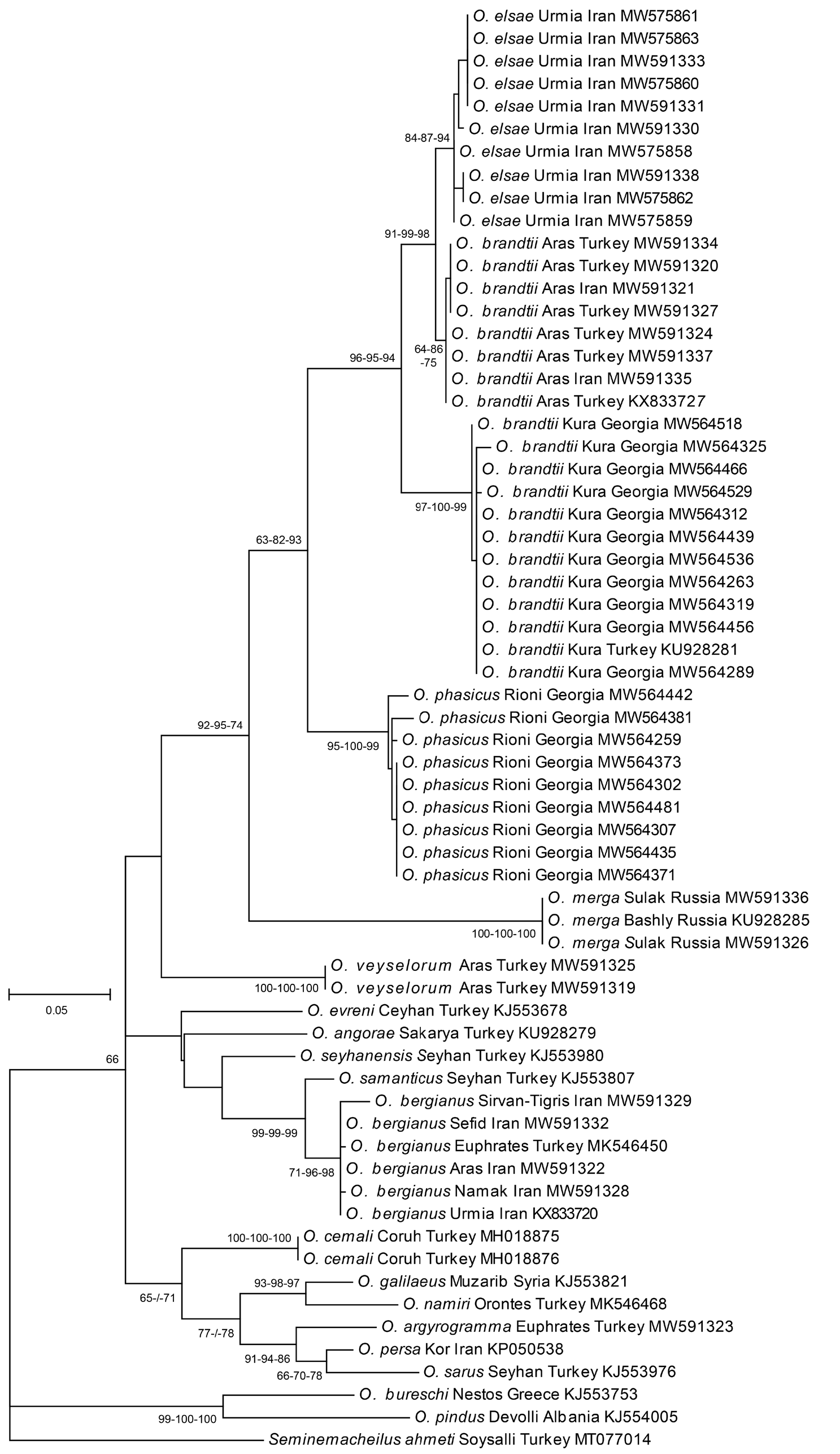
Eagderi et al. (2018) distinguish O. elsae ( Fig. 9 View FIGURE 9 ) from O. brandtii ( Fig. 8 View FIGURE 8 ) based on 10, non-overlapping morphometric characters (dorsal-fin base length, anal-fin base length, anal-fin depth, pelvic-fin length, caudal-fin length, caudal-peduncle width, snout length, eye diameter, interorbital distance, maxillary barbel length). They compared 20 O. elsae with five O. brandtii from the Kura drainage. We examined 20 individuals from each species and found all the 10 morphometric characters overlapping. In seven out of 10 characters no significant difference from equality of their mean values were detected. Only the means of the pelvic-fin length, length of the lower caudal-fin lobe, and the eye diameter were significantly different ( Fig. 10 View FIGURE 10 ). Also these morphometric characters largely overlap: pelvic-fin length 14–20, mean 18% SL in O. brandtii (vs. 14–17, mean 16 in O. elsae ), length of lower caudal-fin lobe 20–27, mean 24% SL (vs. 19–23, mean 21), and the eye diameter 13–21, mean 17% HL (vs. 17–24, mean 20). The overlap in the pelvic-fin length is due to two individuals only and from 49 O. brandtii all except two individuals, have a pelvic-fin length 17–20 % SL, one has 14% SL and one has 15% SL. This character allows to distinguish O. brandtii from O. elsae if a larger number of individuals is available, while the other characters can just support their diagnosis. Oxynoemacheilus elsae is well distinguished from O. brandtii by the shape of the caudal fin, which is emarginate in O. elsae (vs. forked in O. brandtii ) and by details in colour pattern. Oxynoemacheilus elsae usually possess a flank pattern of irregularly shaped and spaced, dark-brown bars, often split in middle, more narrow than interspaces, or flank almost plain brown (vs. flank with brown bars or vertically elongate blotches, much wider than interspaces, usually regularly set) and the caudal fin possess many small brown blotches on rays arranged in 3–6, narrow, pale-brown band (vs. caudal fin with 1–3, wide and very bold bands). However, the flank pattern is variable in both species and occasionally almost plain brown individuals or individuals with many, very narrow bars occur. Oxynoemacheilus elsae is separated from O. brandtii by a minimum K2P distance of 1.5% (Aras) and 4.9% (Kura) in the mtDNA COI barcode region ( Fig. 1 View FIGURE 1 ).
Oxynoemacheilus from the Aras River ( Fig. 11 View FIGURE 11 ), a major tributary to the Kura, cluster as sister to O. elsae in our molecular analysis based on all three approaches (ML, NJ, MP). In all morphological characters distinguishing O. elsae from O. brandtii , the population from the Aras agrees with O. brandtii and not with O. elsae ( Fig. 9 View FIGURE 9 ) including the characteristic colour pattern of O. brandtii . These fish are significantly different from O. elsae in the pelvic-fin length mentioned above, but not different from O. brandtii in any of these or other characters examined. The obvious mismatch between morphological and molecular characters, which suggests that the fish from the Aras should be identified as O. elsae cannot be resolved here. It could be speculated that past river-captures might have resulted in an invasion of a mitochondrial haplotype of O. elsae from Urmia into the adjacent Aras. While they hybridised with native O. brandtii , their mitochondria might have performed better and gave the carriers an evolutionary benefit which spread in the Aras. If so, what cannot be excluded, we would find O. brandtii with mtDNA of O. elsae . The small molecular distance between O. elsae and O. brandtii from the Aras in their mtDNA indicates the time passed since the connection of the Aras with the Urmia basin. Further nuclear molecular research is needed in the future aiming to clarify the case. We suggest to identify the fish from the Aras as O. brandtii due to its morphological similarity to this species.









| ZFMK |
Zoologisches Forschungsmuseum Alexander Koenig |
| MW |
Museum Wasmann |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |