
Athanas exilis, Komai & Henmi, 2023
|
publication ID |
https://doi.org/ 10.11646/zootaxa.5277.2.6 |
|
publication LSID |
lsid:zoobank.org:pub:BD8D27A6-0246-44F7-852C-CB0CDDBBAE8F |
|
DOI |
https://doi.org/10.5281/zenodo.7893015 |
|
persistent identifier |
https://treatment.plazi.org/id/98781E1A-A4A2-499C-8DC8-329D975A1EA6 |
|
taxon LSID |
lsid:zoobank.org:act:98781E1A-A4A2-499C-8DC8-329D975A1EA6 |
|
treatment provided by |
Plazi |
|
scientific name |
Athanas exilis |
| status |
sp. nov. |
Athanas exilis n. sp.
[New Japanese name: Wakasa-murasaki-ebi]
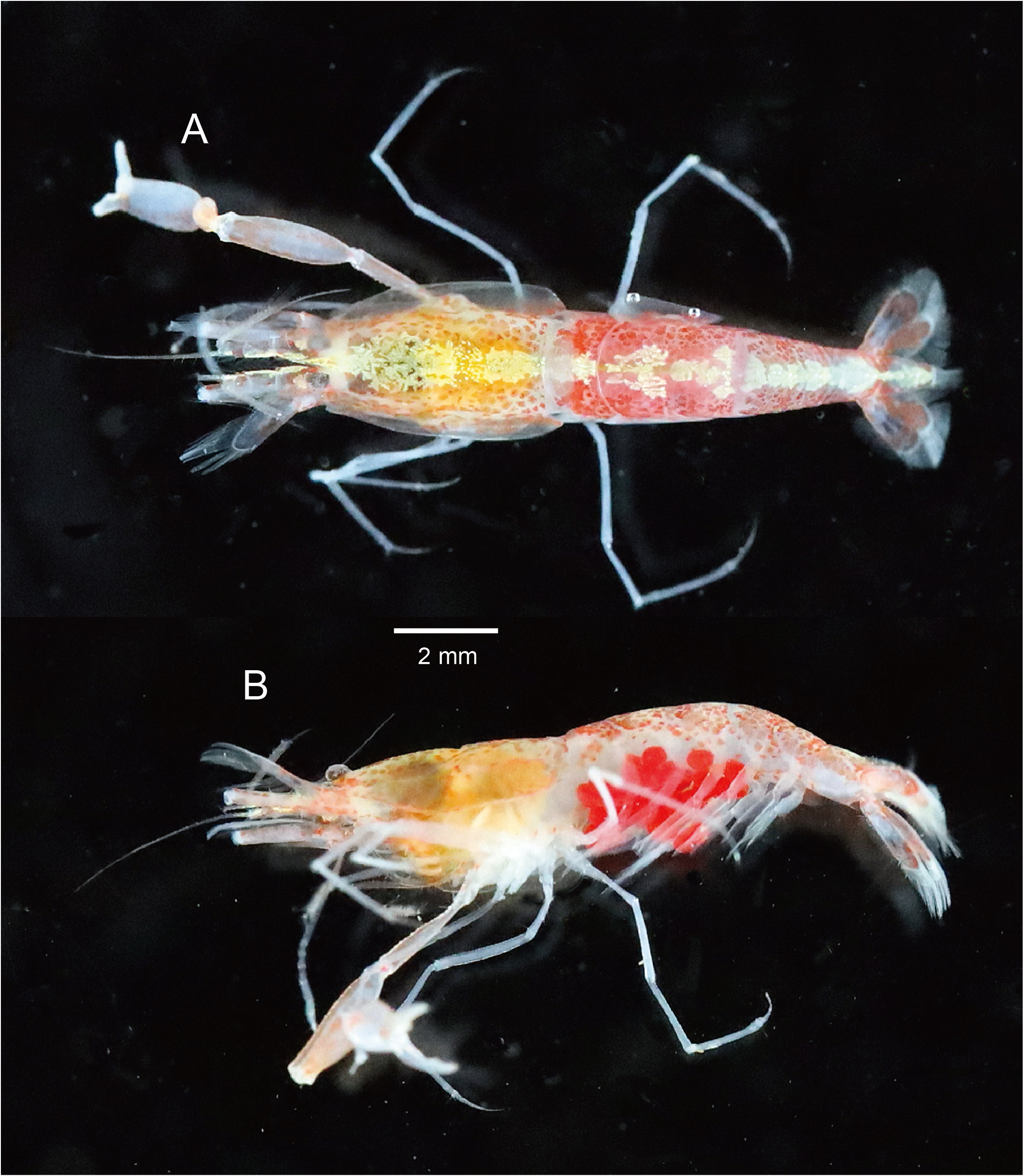
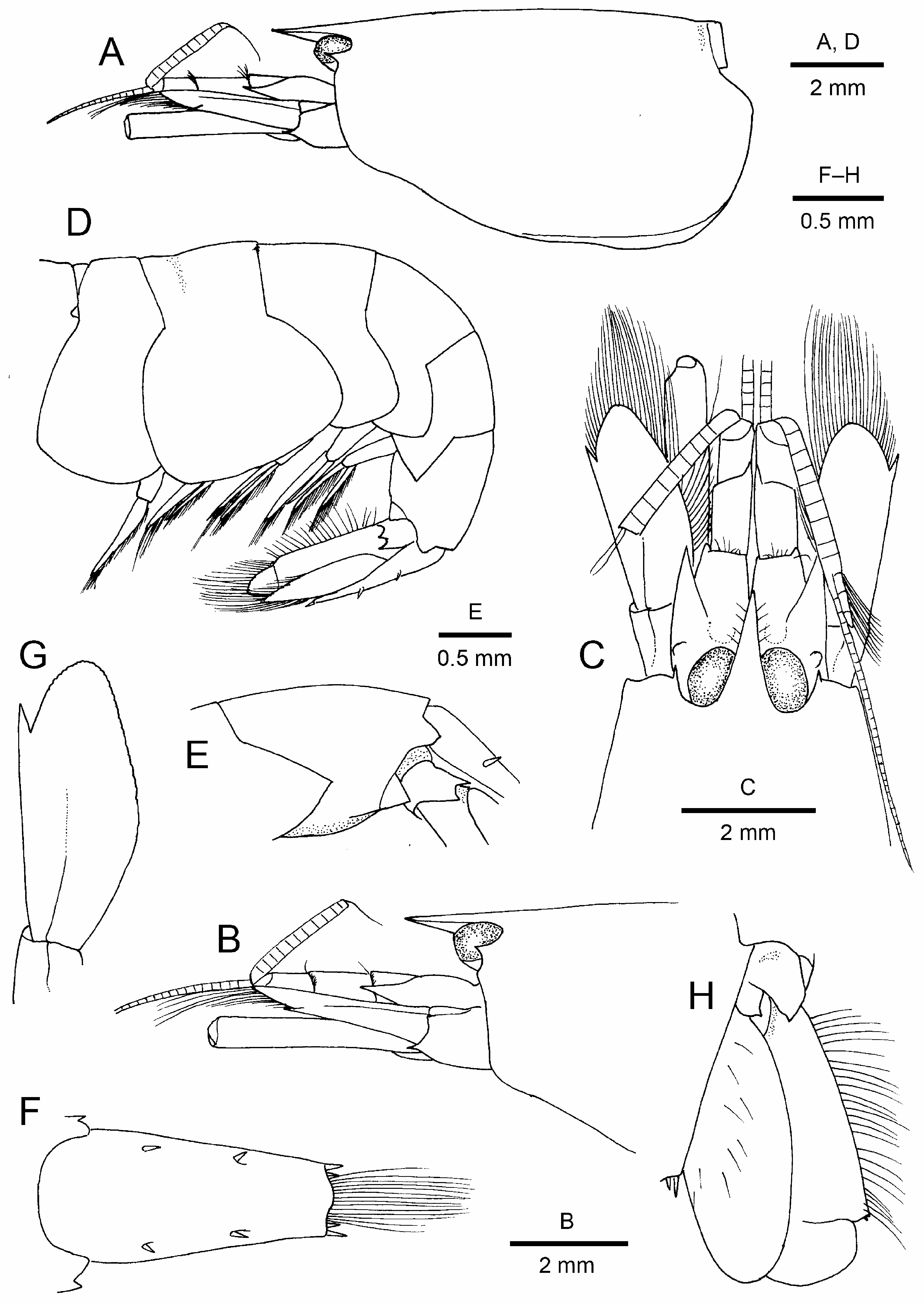
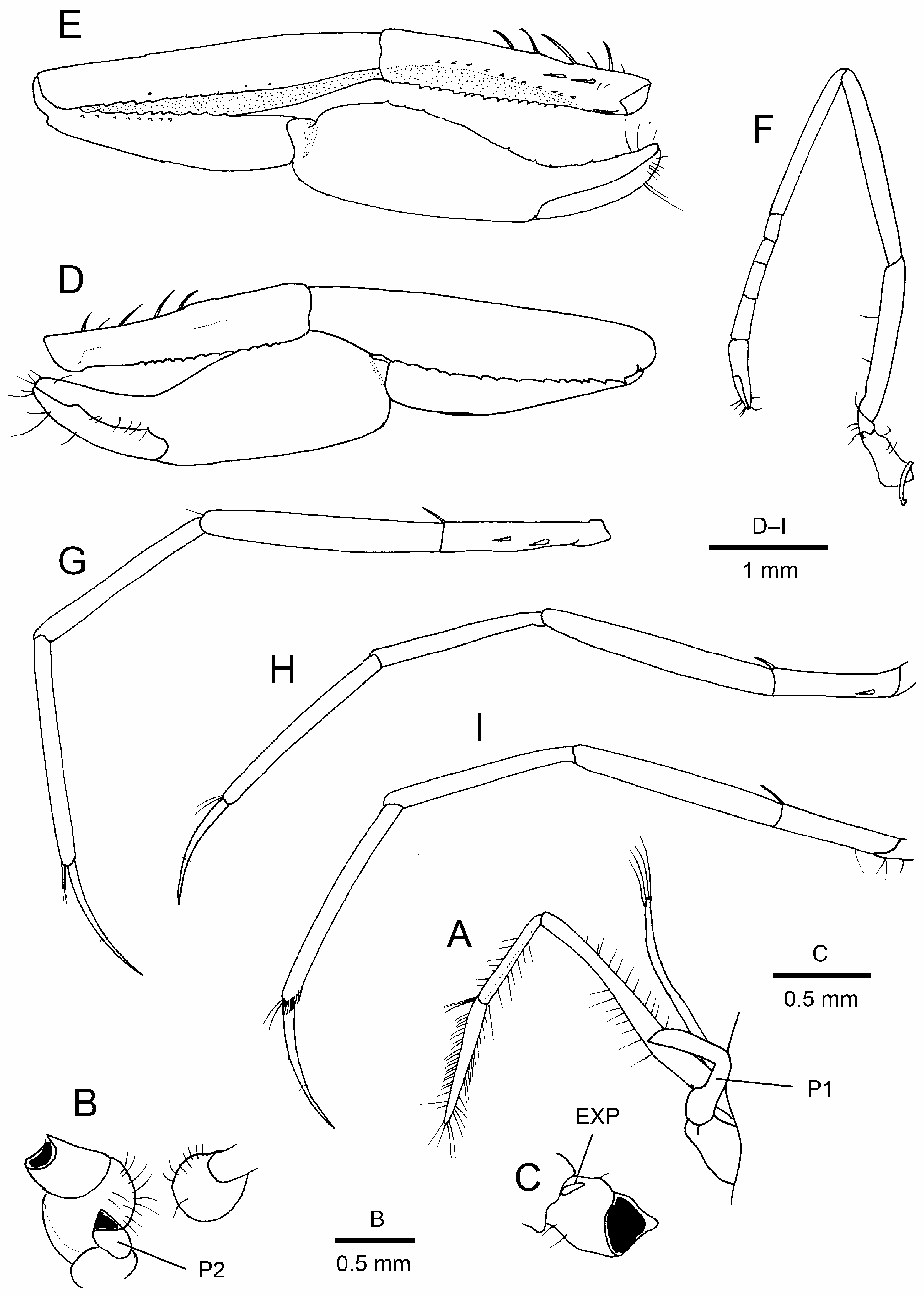
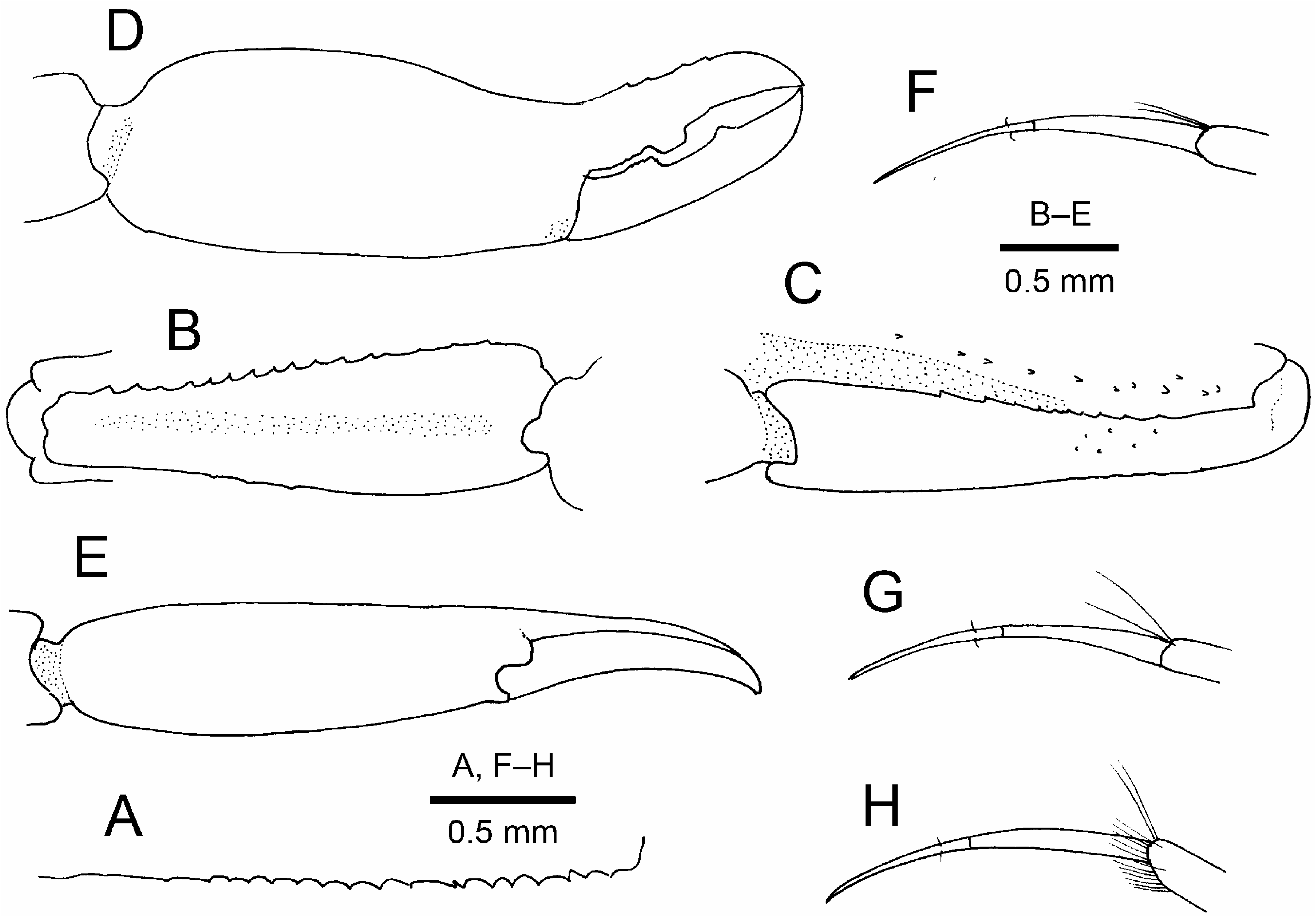
( Figs. 1–4 View FIGURE 1 View FIGURE 2 View FIGURE 3 View FIGURE 4 )
Material examined. Holotype: CBM-ZC 17112, ovigerous female (cl 4.8 mm), Wakasa Bay , NE of Ine, 35°44'47"N 135°20'10”E, 90 m deep, mud, 14 March 2022, RV “Ryokuyo-maru”, SM grab, coll. Y. Henmi. GoogleMaps
Description. Body ( Figs. 1 View FIGURE 1 , 2A, B View FIGURE 2 ) stout.
Carapace ( Fig. 2A–C View FIGURE 2 ) glabrous, not setose. Rostrum 0.2 times as long as carapace, flattened dorsoventrally, directed forward, reaching beyond mid-length of basal article of antennular peduncle, proximally broadened, narrow triangular with acute apex in dorsal view; rostral dorsal carina absent. Frontal margin devoid of supracorneal spines; extracorneal spines acuminate, far falling short of distal corneal margin; infra-corneal spines reduced into low convexity. Pterygostomial margin broadly rounded, not protruding anteriorly. Cardiac notch deep.
Pleura of pleomeres 1–4 ( Fig. 2D View FIGURE 2 ) rounded, unarmed, while that pleomere 5 ( Fig. 2D, E View FIGURE 2 ) acutely pointed at posteroventral angle. Pleomere 6 ( Fig. 2D, E View FIGURE 2 ) with articulated triangular flap posteroventrally; posterolateral process pointed but not acuminate. Telson ( Fig. 2F View FIGURE 2 ) widest at proximal one-fifth, tapering distally, with 2 pairs of dorsolateral spiniform setae; posterior margin medially slightly convex, with 2 pairs of posterolateral spiniform setae, mesial seta almost twice as long as lateral seta.
Eyes ( Fig. 2B, C View FIGURE 2 ) largely exposed in dorsal and lateral views; cornea well pigmented.
Antennular peduncle ( Fig. 2B, C View FIGURE 2 ) basal article longer than distal two articles combined, article 2 slightly longer than article 3. Basal article with stylocerite acute, reaching its distal margin; ventromesial carina with well developed, acute spine. Lateral flagellum biramous, fused proximal portion composed of 8 segments, aesthetasc-bearing shorter ramus (left side damaged) composed of 3 segments.
Antenna ( Fig. 2B, C, G View FIGURE 2 ) with basicerite bearing small ventrolateral distal spine. Scaphocerite reaching distal end of antennular peduncle, oval, lateral margin straight, distal lamella produced, rounded, distinctly overreaching distolateral spine. Carpocerite exceeding distal margin of scaphocerite by 0.2 length, reaching far beyond distal margin of antennular peduncle.
Mouthparts without specific features, typical for genus.
Maxilliped 3 ( Fig. 3A View FIGURE 3 ) endopod slender, reaching distal margin of scaphocerite by tip of ultimate article. Ultimate article 1.2 times as long as penultimate article, tapering to blunt tip bearing only stiff setae (no spiniform setae). Antepenultimate article slightly sinuous, slightly widened proximally. Coxal plate not strongly produced, rounded. Exopod not reaching distal end of endopod antepenultimate article.
Left pereopod 1 only bud-like, in process of regeneration ( Fig. 3A View FIGURE 3 ). Pereopods 1 coxae ( Fig. 3B View FIGURE 3 ) asymmetrical, right larger than left, suggesting notable asymmetry of pereopods 1. Right pereopod 1 (cheliped) ( Fig. 3E, D View FIGURE 3 , 4B–E View FIGURE 4 ) large, carried flexed, overreaching distal margin of scaphocerite by 0.2 length of merus when extended. Basis ( Fig. 3C View FIGURE 3 ) with rudimentary exopod. Ischium elongate, 0.8 times as long as merus; ventral margin with row of numerous spinules and 1 proximal spiniform seta, not particularly projecting; dorsal margin with row of 4 long, slender spiniform setae on proximal half; ventromesial facet clearly delimited, mesial margin with row of spinules; proximomesial face with 2 long spiniform setae. Merus elongate; dorsal, lateral and mesial surfaces smooth; ventral surface deeply excavate to accommodate carpus and proximal part of chela, ventrolateral ( Fig. 4A View FIGURE 4 ) and ventromesial margins sharply edged, each with row of widely spaced spinules or granules. Carpus 0.8 length of merus, distally widening, somewhat compressed, without setose area; extensor surface with faint median sulcus; lateral surface smooth, unarmed; mesial surface with 2 irregular longitudinal rows of spinules or granules along flexor margin. Chela also somewhat compressed, distinctly wider than carpus, surface glabrous; mesial margin sinuous with concavity at pollex base; pollex deflexed, terminating in minute clow, with low, rectangular tooth on occlusal margin slightly proximal to midlength; dactylus gradually tapering, crossing fixed finger at tip, 0.7 times as long as palm, occlusal margin with small triangular tooth slightly distal to midlength.
Pereopod 2 ( Fig. 3F View FIGURE 3 ) slender, overreaching distal margin of scaphocerite by half-length of carpus, without exopod. Ischium slightly shorter than merus. Carpus composed of 5 segments, first segment longer than four others combined; ratio of carpal segments (from proximal to distal) approximately: 6.2: 1: 1: 1.5: 1.6. Chela simple, longer than distal carpal segment, fingers slightly longer than palm.
Pereopods 3–5 slender, generally similar. Pereopod 3 ( Fig. 3G View FIGURE 3 ) overreaching distal margin of scaphocerite by full length of propodus; ischium with 1 slender spiniform seta at dorsodistal end and 2 spiniform setae on lateral surface ventrally; merus, carpus and propodus unarmed, merus 7.3 times as long as broad; dactylus ( Fig. 4F View FIGURE 4 ) 0.6 times as long as propodus, simple, very slender, gently curved, with paired minute setae slightly distal to midlength. Pereopod 4 ( Figs. 3H View FIGURE 3 , 4G View FIGURE 4 ) overreaching distal margin of scaphocerite by half-length of propodus; ischium with 1 dorsodistal and 1 ventrolateral spiniform setae. Pereopod 5 ( Figs. 3I View FIGURE 3 , 4H View FIGURE 4 ) overreaching distal margin of scaphocerite by length of dactylus; ischium with 1 dorsodistal spiniform seta, but no ventrolateral spiniform seta; propodus with grooming setae distally.
Gill formula typical for genus: pleurobranchs on thoracomeres 4–8 (above pereopods 1–5); podobranch and arthrobranch absent; lobe-like epipods on maxilliped 1 and 2, strap-like epipods on coxae of maxilliped 3 and pereopods 1–3; setobranchs on coxae of pereopods 1–4; maxillipeds 1 and 2 each with well-developed, flagellumlike exopod.
Pleopod 2–5 each with appendix interna on endopod.
Uropod ( Fig. 2H View FIGURE 2 ) with protopod bearing two distally acute lobes. Exopod lateral margin nearly straight, terminating in minute denticle; distolateral spiniform seta minute; diaeresis straight, unarmed. Endopod subovate, subequal in length to exopod.
Colouration in life. Carapace and pleon semitransparent with scattered red dots (sparse on carapace), dorsal surface with yellowish white longitudinal band along midline; yellowish hepatopancreas visible through integument; antennule and antenna semitransparent; cheliped whitish with tinge of pink; pereopods 2–5 whitish semitransparent; uropods pinkish; eggs red ( Fig. 1A, B View FIGURE 1 ).
Habitat. The holotype of the new species was extracted from muddy sediment collected using a Smith McIntyre grab at a depth of 90 m. The bottom salinity was 34.3‰ and the bottom water temperature was 11.8°C on 14 March 2022.
Distribution. Presently known only from Wakasa Bay, the Sea of Japan side of Honshu, Japan, at a depth of 90 m.
Remarks. In the holotype, the known unique representative of the new taxon, the left cheliped is in the process of regeneration ( Fig. 3A View FIGURE 3 ). Nevertheless, the coxae are asymmetrical (right coxa larger than left), indicating that the chelipeds are distinctly unequal and dissimilar. It is assumed that the right cheliped of the holotype corresponds to a major cheliped because of the large size and stoutness.
The important features of Athanas exilis n. sp. for species differentiation are: the rostrum does not reach the distal margin of the basal article of the antennular peduncle, with a smooth, non-dentate dorsal margin; no infracorneal and supracorneal spines are present on the carapace frontal margin; the antennular peduncle is slender, with the article 2 being distinctly longer than wide; the antennal carpocerite overreaches the distal margin of the scaphocerite; the major cheliped is carried folded; the major cheliped ischium and merus have spinulose ventral margins; the major cheliped fingers are not twisted and devoid of setal brush; the major cheliped carpus is elongate, being longer than the palm; and the pereopods 3–5 are slender, with simple, noticeably elongate dactyli exceeding 0.5 length of propodi. This combination of characters is seen only in A. sydnyensis .
Athanas sydnyensis is known only by the type series from New South Wales, Australia, at depths of 4–10 m, consisting of a female holotype and four female paratypes ( Anker & Ahyong 2007). Athanas exilis n. sp. differs from A. sydnyensis in the following particulars: the dorsal surface of the rostrum is devoid of a middorsal carina in A. exilis n. sp., which is discernible in A. sydnyensis (cf. Fig. 2C View FIGURE 2 versus Anker & Ahyong 2007: fig. 1B); the lateral margins of the telson are straight in A. exilis n. sp., rather than gently convex in A. sydnyensis (cf. Fig. 2F View FIGURE 2 versus Anker & Ahyong 2007: fig. 1J); the antennular stylocerite reaches the distal margin of the basal peduncular article in A. exilis n. sp., while reaching the midlength of the peduncular article 2 in A. sydnyensis (cf. Fig. 2C View FIGURE 2 versus Anker & Ahyong 2007: fig. 1A–C); the distal lamella of the antennal scaphocerite is more strongly produced in A. exilis n. sp. than in A. sydnyensis (cf. Fig. 2G View FIGURE 2 versus Anker & Ahyong 2007: fig. 1B, C); the pereopod 2 is much slenderer in A. exilis n. sp. than in A. sydnyensis (cf. Fig. 3F View FIGURE 3 versus Anker & Ahyong 2007: fig. 1H); the ventral armature on the pereopod 1 merus is less developed in A. exilis n. sp. than in A. sydnyensis (cf. Fig. 3D, E View FIGURE 3 versus Anker & Ahyong 2007: figs. 1D–G, 2A, B). In addition, as mentioned above, the chelipeds are probably unequal and dissimilar in the new species, but they are subequal and similar in A. sydnyensis ( Anker & Ahyong 2007: figs. 1D–G, 2A, B).
Besides the development of the infracorneal spine on the carapace, Athanas amazone is also similar to the new species. In A. amazone , the infracorneal spine is usually present, but occasionally absent ( Anker & Ahyong 2007). Athanas amazone has been reported from the eastern Atlantic, i.e., Nigeria ( Holthuis 1951), Togo, Congo ( Crosnier & Forest 1973), Cape Verde (Türkay 1982), Morocco ( García Raso 1996), and throughout the Mediterranean Sea ( Holthuis & Gottlieb 1958; Crosnier & Forest 1973; Kocataş 1981; Koukouras et al. 1992; Froglia & Argenti 1993; García Raso 1996; Atkinson et al. 1997; Froglia & Atkinson 1998; d’Udekem d’Acoz 1999; Anker &Ahyong 2007). Holthuis & Gottlieb (1958), Crosnier & Forest (1973) and Anker & Ahyong (2007) noted some variation in the relative length of the stylocerite (compared to the distal margin of the first segment of the antennular peduncle); the length of the carpocerite (compared to the scaphocerite); and the shape of the chelipeds. Anker & Ahyong (2007) stated: “It shows that some features, e.g., the length of the carpocerite, the shape of the merus and the chela of the major cheliped, and the slenderness of the minor cheliped, may indeed be sexually dimorphic and/or age-dependent. However, more material with information on colour patterns and ecology is needed to conclude about the taxonomic status of A. amazone .” Considering the possible sexual dimorphism and the possibility that more than one species might be confounded under A. amazone , the following comparison is limited to the accounts mentioning females ( Holthuis 1951; Anker & Ahyong 2007). Athanas exilis n. sp. differs from A. amazone as follows: the distal lamella of the antennal scaphocerite is more strongly produced in A. exilis n. sp. than in A. amazone ( Fig. 2G View FIGURE 2 versus Holthuis 1951: fig. 23c); the major cheliped merus is less expanded in A. exilis n. sp. than in A. amazone ( Fig. 3D, E View FIGURE 3 versus Holthuis 1951: fig. 23e); the pereopod 2 is much more slenderer in A. exilis n. sp. than in A. amazone ( Fig. 3F View FIGURE 3 versus Holthuis 1951: fig. 23g); the major chela is longer than the carpus in A. exilis n. sp., whereas shorter than the carpus in A. amazone ( Fig. 3D, E View FIGURE 3 versus Holthuis 1951: 23 e).
A partial fragment of the 16S rRNA gene (542 bp) was sequenced for the holotype of the present new species. Sequences of the16S rRNA gene are available for eight species (nine sequences) of Athanas in the GenBank ( Table 1 View TABLE 1 ), although no sequence data is available for A. sydnyensis and A. amazone , mentioned above. The interspecific K2P genetic divergence among the nine Athanas species is high, ranging from 14.0 to 40.4% ( Table 2 View TABLE 2 ). The genetic divergence between Athanas exilis n. sp. and the other eight species ranges from 21.1 to 33.4%. It should be noted that the identity of the voucher specimen of the sequence KU312972 with A. nitescens Leach, 1813 is questionable, because the sequence closely matches MZ661664 referred to A. glimardii Coutiére, 1911 (K2P genetic divergence 0.8%), rather than to MZ661665 referred to A. nitescens (K2P genetic divergence 14.8%).
Of the 13 species known from Japan, 10 species are widely distributed in the tropical or subtropical Indo-West Pacific ( Miya & Miyake 1968; Banner & Banner 1973; Chace 1988; Hayashi 1995a; Sha et al. 2019), whereas three species, including the present new species, are so far restricted to temperate areas in East Asian waters: A. squillophilus was originally described from Ishikari Bay, Hokkaido, Japan ( Hayashi 2002), and later recorded from the Yellow Sea coast of northern China ( Sha et al. 2019); A. alpheusophilus Marin, 2017 was originally described from Posjeta Bay, Far Eastern Russia (Sea of Japan); and A. exilis n. sp. is so far known only from Wakasa Bay, Japan, although its real distribution remains clarified. The holotype of A. exilis n. sp. was collected from the sea bottom of the temperature 11.8°C (see above), suggesting that the new species is adapted to the temperate environment.
Etymology. From the Latin exilis (= slender) in reference to the slender, elongate dactyli of the pereopods 3–5 in the new species.
| RV |
Collection of Leptospira Strains |
| SM |
Sarawak Museum |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |