
Lepidiolamprologus mimicus, Schelly, Robert, Takahashi, Tetsumi, Bills, Roger & Hori, Michio, 2007
|
publication ID |
https://doi.org/ 10.5281/zenodo.179611 |
|
DOI |
https://doi.org/10.5281/zenodo.5681449 |
|
persistent identifier |
https://treatment.plazi.org/id/03AE4E67-FF92-FFEF-FF24-F988FB0B164E |
|
treatment provided by |
Plazi |
|
scientific name |
Lepidiolamprologus mimicus |
| status |
sp. nov. |
Lepidiolamprologus mimicus View in CoL new species
Figs. 1–5 View FIGURE 1 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 , Table 1 View TABLE 1
Holotype: SAIAB 41257, Chituta Bay, Zambia, at Chituta Bay cliffs, at 10–15 m depth along the rocky slope, Nov. 17, 1992, R. Bills. (approximate coordinates: 08° 43.82 S, 31° 09.41 E)
Paratypes: SAIAB 58308 (n=1; cleared and stained), collected with holotype; FAKU 95137 (male, 123.1 mm SL), FAKU 95138 (female, 121.7 mm SL), Kasenga, Zambia, 14 m depth, 7 Nov. 2005, gill net, M. Hori. FAKU 95139 (male, 159.7 mm SL), Kasenga, Zambia, 13–16 m depth, 3 Dec. 2005, gill net, M. Hori. FAKU 95140 (male, 119.4 mm SL), Kasenga, Zambia, 2–15 m depth, 30 Oct. 1996, gill net, M. Hori. FAKU 95141 (male, 129.6 mm SL), Kasenga, Zambia, 15 m depth, 12 Dec. 1996, gill net, H. Ochi. FAKU 95142 (male, 76.9 mm SL), Kasenga, Zambia, 19 m depth, 1 Nov. 1995, M. Hori. FAKU 95143 (male, 101.7 mm SL), off Mtondwe Is., Zambia, depth unknown, 1 Nov. 1995, gill net, M. Hori. FAKU 95144 (female, 133.6 mm SL), Kasenga, Zambia, 14–19 m depth, 13 Nov. 1995, gill net, M. Hori. FAKU 95145 (male, 147.0 mm SL), FAKU 95146 (female, 130.7 mm SL), Kasenge, Zambia, 15 m depth, 11 Jan. 1994, gill net, M. Hori.
Differential diagnosis. From the Lepidiolamprologus sensu Poll (1986) , Lepidiolamprologus mimicus n. sp. is distinguished from L. cunningtoni by, among numerous other neurocranial and gill arch characters, possession of a sesamoid bone in the labial ligament, 2 pores at NLF0, and a color pattern based on three rows of irregular spots, whereas L. cunningtoni lacks a labial ossification, has a single, coalesced NLF0 pore, and a color pattern based on a series of faint bars. With the other five members of Poll’s Lepidiolamprologus , L. mimicus n. sp. shares a whole suite of neurocranial, gill arch, and coloration characters, but is still easily distinguishable. Of these species, it is unique in having bright yellow fins and very dark, large spots on the flanks, without additional small spots or a complex pattern of spots and worm lines on the head (as in L. nkambae and L. kendalli ). Lepidiolamprologus mimicus n. sp. is somewhat deeper-bodied than L. nkambae , L. kendalli , and L. attenuatus (BD 23.4–26.2% SL, vs. max 23.3% SL in L. nkambae ), has a more elongate head than L. attenuatus (HL 31.0–33.7% SL, vs. 28.2–30.5% SL in L. attenuatus ), has one more vertebra than both L. nkambae and L. kendalli (34 vs. 33), and has a wider interorbital region (IW 17.4–24.5% HL) than L. nkambae (IW 14.3–16.9% HL) and L. kendalli (IW 14.3–17.8% HL). In addition, it usually has one more dorsal fin ray than both L. nkambae and L. kendalli (11 vs. 10). The longitudinal line scale count of L. mimicus n. sp.
is almost always higher (73–79) than in L. attenuatus and L. elongatus (both 66–73), L. kendalli (67–70) and L. profundicola (66–74). Finally, L. mimicus n. sp. generally has fewer gill rakers (10–12) on the first ceratobranchial than L. elongatus (11–14) and L. attenuatus (13–15).
Description. Counts and measurements for the holotype and 12 paratypes given in Table 1 View TABLE 1 . An elongate, moderately shallow-bodied species (BD 23.4–26.2, mean 24.7% SL), with a large mouth and sleek form characteristic of an open-water piscivore. Greatest body depth at about base of fifth dorsal fin spine. Head length 31.0–33.7, mean 32.4% SL. Head profile slightly convex, rising smoothly to dorsal fin base.
Fins. Dorsal fin XVIII 10–11, anal fin V 8–9. Spines in the dorsal fin increase gradually in length posteriorly until about the 7th spine, after which they remain constant. Anal fin spines gradually increase posteriorly through entire series. Dorsal and anal fins come to a point and terminate at about the end of the caudal peduncle; no rays are produced. Caudal fin large and emarginate, with 14 branched rays. Pectoral and pelvic fins short, terminating well ahead of anus. First ray in pelvic fin is longest.
Jaws and Teeth ( Fig. 2 View FIGURE 2 ). Lower jaw prognathous, with both outer and inner row teeth pointed unicuspids in both jaws. Single series of enlarged, recurved, procumbent caniniform teeth (8 premaxillary and 6 dentary) situated on anterior third of both jaws, with largest teeth furthest from symphysis. Posterior to large teeth single rows of slightly enlarged caniniform teeth run almost whole length of dentigerous arms of both dentary and premaxilla, outer row on premaxilla with 49–60 teeth. Small, caniniform inner teeth in five to nine irregular rows nearest symphysis, tapering to single row posteriorly and running most of length of dentigerous arms of premaxilla and dentary.
Total lateral line scales 75 73 (1), 74 (2), 75 (3), 76 (2), 77 (2), 78 (1), 79 (2) Upper branch lateral line 52 51 (1), 52 (1), 53 (2), 54 (1), 56 (1), 57 (1), 58 (1), 60 (1), 63 (3), 67 (1) Lower branch lateral line 37 26 (2), 29 (1), 30 (1), 31 (2), 32 (1), 34 (2), 36 (1), 37 (1), 40 (1), 45 (1) Dorsal fin spines and rays XVIII 11 XVIII 10 (2), XVIII 11 (11) Anal fin spines and rays V 9 V 8 (5), V 9 (8)
Gill rakers—lower arm of 1st arch 10 10 (5), 11 (5), 12 (3)
Vertebrae 16+18 16+18 (n=3)
Gill rakers ( Fig. 3 View FIGURE 3 a). Slender, elongate, and non-denticulate, with up to four subsidiary branches. Thirteen to 16 gill rakers along outer row of first gill arch. No rakers present on hypobranchial, 10–12 rakers along ceratobranchial, sometimes single raker in angle of arch, and 3–4 rakers on epibranchial.
Lower pharyngeal jaw ( Fig. 3 View FIGURE 3 c, d). Longer than wide, with slight interdigitation along ventral suture. About 24 teeth in posterior tooth row. All teeth slender and beveled or hooked.
Scales. Flank scales small, ctenoid, and regularly imbricating. Lateral line scales 73–79, with 51–67 in upper and 26–45 in lower lateral line branch. Considerable overlap (16–20 scales) between upper and lower branches of lateral line. Cheek usually naked, but 4–5 cheek scales observed in three of 13 individuals, small scales on belly and small, deeply embedded scales on chest. Opercle and subopercle heavily scaled. Caudal fin with scales extending more than half its length.
Vertebrae. 34; 16+18.
Additional osteology ( Figs. 3 View FIGURE 3 , 4 View FIGURE 4 ). Infraorbital series comprised of broad, plate-like lachrymal with 6 large sensory canal pores, lacking infraorbitals adjacent to lachrymal. Dermosphenotic present. Labial ossification present. Single supraneural present. Supraoccipital crest well-developed and increasing in depth posteriorly, with sharply-angled posterior edge. No frontal ridge present, two distinct pores at NLF0. Frontal shelf welldeveloped and extending anteriorly beyond NLF2. Pre-orbital process of lateral ethmoid rotated posteromedially. Post-orbital process of sphenotic followed by a large, spoon-shaped wing. Frontal and ethmoidvomer continuous in profile, without abrupt drop-off.
Coloration. In life, background body color brownish-tan, with three broken white to silvery-blue stripes composed of irregular blotchy spots. Superimposed over this pattern are three rows of seven to nine large, dark-brown spots stretching from above opercle to caudal flexure. Large spots irregular, with intervening space between them horizontally about as large as the spots. Vertically, some spots merge together partially, though some are staggered irregularly resulting in misalignment between horizontal rows. Largest spots in lower row, which is superimposed over the lower branch of lateral line at about center of flank, while middle row of slightly smaller spots is superimposed over upper lateral line branch. Top row of spots sits along dorsal fin base, with spots stretching partially onto dorsal fin. Opercle with a patch of lavender and a black blotch posteriorly, belly white, eyes with bright yellow dorsal and ventral bordering. Bluish-white, thin line running below eye from lachrymal to whitish patch on cheek. All fins with bright yellow; dorsal, anal, and caudal fins with rows of large yellow maculae interspersed with small patches of white. Yellow maculae blend together vertically to form series of yellow bands. Pectoral fin pale yellow on the base. Preserved coloration a uniform, light yellow-tan with brown spots in pattern described above. No sexual dimorphism in coloration.
Diet. Rocky littoral, open-water piscivore.
Ecology and behavior. ( Fig. 5 View FIGURE 5 ). Ecology and behavior of the holotype and paratypes was not recorded. However, these aspects of the biology of Lepidiolamprologus mimicus n. sp. were studied at Kasenga from 1993 to 2006. Underwater observation during the period and analysis of stomach contents of samples revealed that this species was exclusively piscivorous and mainly preyed on young and sub-adults of cyprichromine cichlids. In the littoral region of Kasenga, four species of cyprichromine ( Cyprichromis leptosoma , C. zonatus , C. coloratus , and Paracyprichromis brieni ) form mixed-species schools composed of various subgroups of age or sex of each species in open water 1–4 m above the rocky substrate ( Takahashi et al. 2002, 2006). Usually L. mimicus n. sp. stalked solitarily near or mingled in the school at a depth of 10 m or more, and attacked mainly young and juveniles of the school. At least 30–40 individual hunts were observed. When hunting, adults of L. mimicus n. sp. changed their body coloration markedly from dark-brown to pale beige; their characteristic dark-brown spots disappeared and rows of fine silver dots on the flank faded, but the yellow anal fin and black dotted line at the tip of the dorsal fin remained. This coloration generally looked like that of female cyprichromines and specifically very much resembled that of female Paracyprichromis brieni . The degree of resemblance in coloration of L. mimicus n. sp. to its prey during hunting was greater in young and sub-adults than adults ( Fig. 5–3 View FIGURE 5 , -4), and the predominant prey were young and juveniles of Paracyprichromis brieni , with small Cyprichromis making up the balance. These phenomena indicate both that L. mimicus n. sp. is highly specialized to forage on cyprichromine fishes and also that its coloration, especially Collection localities of Lepidiolamprologus mimicus Holotype locality represented by open circle.
Mating and breeding of L. mimicus n. sp. have not been observed despite the long-term underwater survey from 1993 to 2006. However, on two occasions, pairs were observed roaming together for several days around the same area at a depth of 20– 25 m. In both cases, the pairs were wandering around a boundary between rocky and sandy substrate, and sometimes inspected holes or crevices among boulders, but did little foraging.
After the pairs were caught by gill-net underwater, gentle pressure on their abdomens made the males eject semen and females eject mature eggs. These observations suggest that L. mimicus n. sp. is a monogamous species like L. elongatus and L. attenuatus , and not a harem species like L. profundicola (for breeding habit of Lepidiolamprologus , see Kuwamura 1997). It also seems that they breed in rather deep water (more than 30 m) beyond the reach of a diving survey. Mature males and females were usually caught with gill-nets in deep water from 20 m to 50 m depth.
Distribution. (Fig. 6). Zambian coast of Lake Tanganyika from Kasenga to Kapembwa. The west side of Chituta Bay, the type locality, is characterized by a steep rocky shore starting in the south at the west side of the mouth of the Lunzua River, with usually poor visibility (<10 m). From the river mouth, where the rocks run to about 5 m depth, the depth of the rocky slope increases steadily until the point at Kasenga village where it is approximately 50 m deep. Specimens of Lepidiolamprologus mimicus n. sp. were collected as solitary individuals wandering across the steep sloped rocky habitat, comprised of large rocks and rock slabs (some several cubic meters). Other Lepidiolamprologus occurring sympatrically in Chituta Bay are L. cunningtoni , L. kendalli , L. elongatus , L. attenuatus , and L. profundicola . Only L. nkambae is absent. According to Konings (1988; page 188 and photo on page 121), Lepidiolamprologus mimicus n. sp. also occurs in central Tanzania.
Etymology. The specific name, mimicus , is from the Latin, in reference to the specific feeding ecology involving imitative coloration described herein.
Remarks. Lepidiolamprologus mimicus n. sp. exhibits all of the features that Poll (1986) used to characterize Lepidiolamprologus : a high number of ctenoid scales (>60) in the longitudinal line; more than 20 scales in the lower lateral line; dorsal fin formula XVII–XXI, 10–12; anal fin formula IV–V, 7–9; and large size (140–305 mm TL). In Poll’s (1986) classification, Lepidiolamprologus included the following elongate, predatory species: L. elongatus , L. attenuatus , L. profundicola , L. kendalli , L. nkambae , and L. cunningtoni . To the exclusion of L. cunningtoni , L. mimicus n. sp. and the remaining five species of Poll’s Lepidiolamprologus all possess an ossification in the labial ligament, two distinct pores at NLF0, forked or stellate gill rakers, scales on the caudal fin to approximately 80% of caudal length, and three series of spots along the body, situated at the mid-line, upper lateral line, and dorsal fin base.
Mimicry in its various forms is fairly widespread in marine fishes ( Randall 2005; Moland et al. 2005). A common form is aggressive mimicry, in which a predatory species resembles a harmless species, allowing it to avoid detection when approaching prey ( Wickler 1968). In addition to the numerous marine examples, several tropical freshwater characids, citharinids, and cichlids have been recognized as aggressive mimics ( Sazima 2002). Within Lake Tanganyika, aggressive mimicry has been described in scale-eating perissodines ( Yanagisawa et al. 1990; Hori & Watanabe 2000), but this is the first case observed in lamprologines. Interestingly, the abundant schools of brightly-colored Lake Tanganyika cyprichromines are subject to aggressive mimicry from two independent cichlid lineages. Color morphs of Perissodus microlepis with yellow anal fins insinuate themselves into schools of cyprichromines to feed on scales, while yellow-finned Lepidiolamprologus mimicus n. sp. blend into schools to pick off whole fish.
Comparative materials.
Lepidiolamprologus attenuatus : AMNH 98043 (alcohol and 1 C&S); MCZ 49254 (alcohol and 1 C&S); SAIAB 41253; Zm 031-a, 031-b, 0 5018, 05097-2, 0 5193, 25041, 840-a.
Lepidiolamprologus cunningtoni : SAIAB 42470 (alcohol and 1 C&S); MCZ 32596 (1 C&S); Zm 959- 1~8.
Lepidiolamprologus elongatus : AMNH 97219; SAIAB 44646 (alcohol and 1 C&S); Zm 04049-2~3, 04085-1, 04085-3, 04284-1~2, 04284-4~5, 0 4305.
Lepidiolamprologus kendalli : MRAC 77-36-P-1 (paratype, male, 129.9 mm SL), N. W. of Mutondwe Isl., Zambia, 40 m depth, 16 Nov. 1972; SAIAB 46848 (1 alc); SAIAB 46874 (2 alc); Zm 04088-1~2, 04111-1~2, 0 4188, 0 4306, 04621-1~2, 0 4636.
Lepidiolamprologus mimicus n. sp.: AMNH 237601 (n=1; tissue taken), Kapembwa, Zambia, 08°37.37'S, 30°50.87'E, Mar. 10, 2004, R.Bills and R. Schelly.
Lepidiolamprologus nkambae : MRAC 77-49-P-1 (holotype, female, 113.0 mm SL), Nkamba bay, Sumbu National Park, Zambia, Apr. 1975; AMNH 216075 SW (9 C&S); Zm 04658-1~3, 05696-1~2, 97651-a~b, 98138.
Lepidiolamprologus profundicola : MRAC 114.160 (holotype, female, 211.3 mm SL), 114.161 (paratype, female, 222.2 mm SL), Stat: 68, Baie au Sud immidiat du cap Tembwe, Congo, 15 Jan. 1947; MRAC 114.162 (paratype, male, 246.9 mm SL), Stat: 140, Autour de l’ile de Kabimba, 23 Feb. 1947; MRAC 82.12.P.518-519 (1 C&S); IRSNB 345 (paratype, male, 167.9 mm SL), collected with holotype; SAIAB 41255; Zm 0 4004, 0 4050, 0 4402, 0 4463, 0 4533, 04560-1, 0 4565, 04570-1, 04597-1~2, 04622-1, 04629-2.


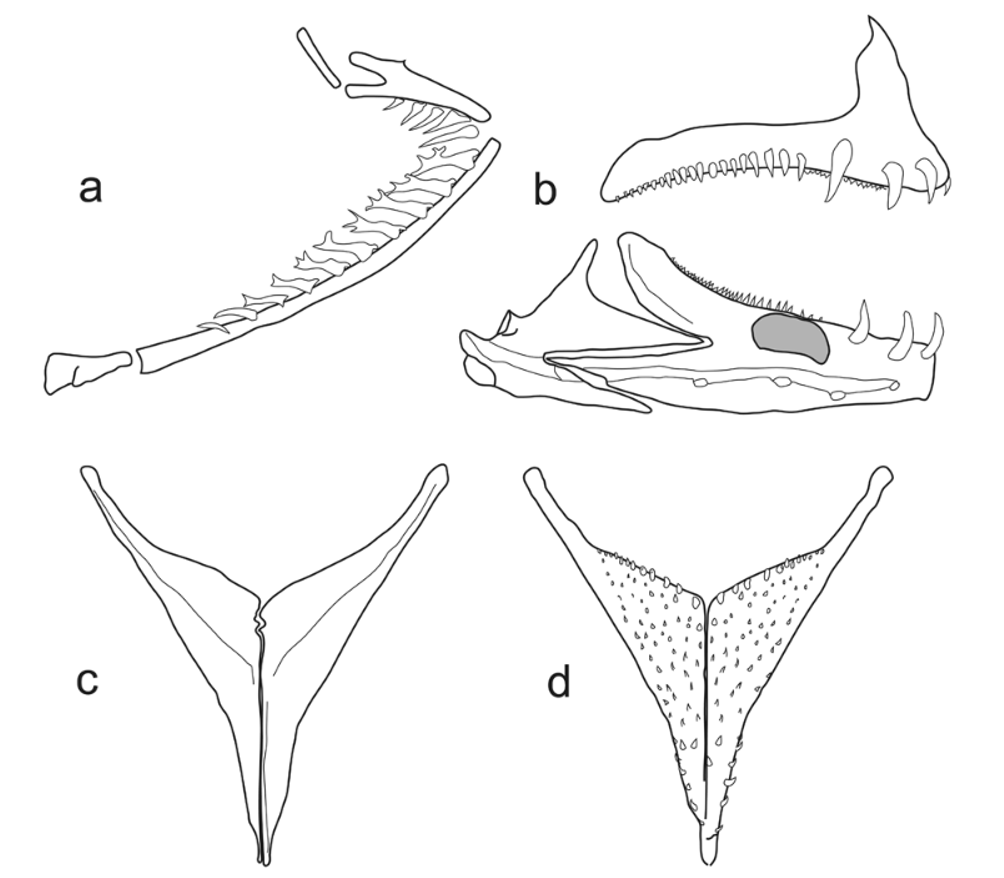
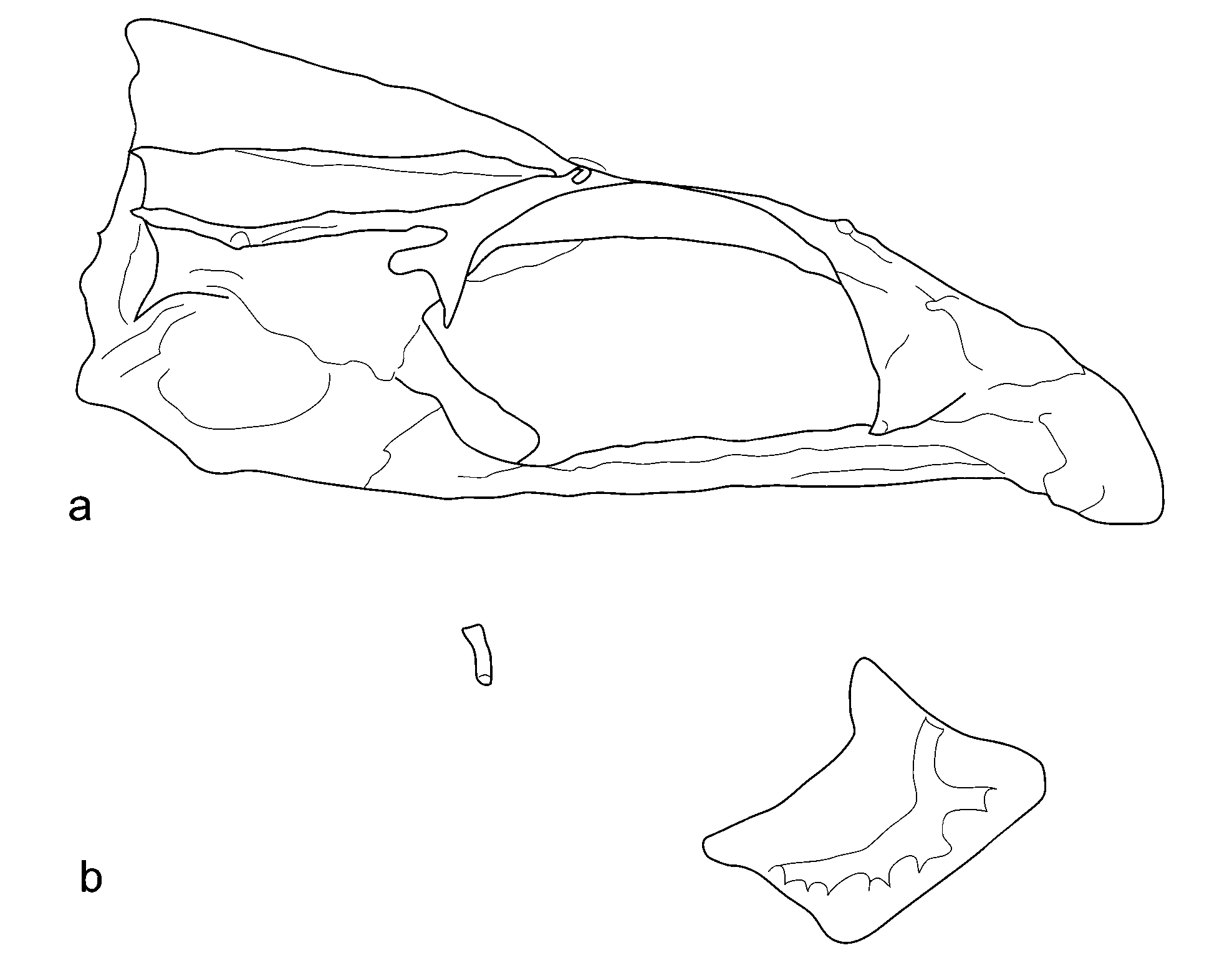

TABLE 1. Morphometric and Meristic data for Lepidiolamprologus mimicus n. sp.
| Character | holotype n | mean | min | max | SD |
|---|---|---|---|---|---|
| Standard length (mm) | 144.8 13 | 121.3 | 76.9 | 159.7 | 23.8 |
| Percent of standard length | |||||
| Body depth Head length | 25.4 13 33.1 13 | 24.7 32.4 | 23.4 31.0 | 26.2 33.7 | 0.98 0.69 |
| Caudal peduncle depth | 10.2 12 | 10.4 | 10.1 | 10.7 | 0.19 |
| Caudal peduncle length Anal fin base length | 14.0 13 20.1 13 | 16.4 19.8 | 14.0 17.7 | 19.4 21.4 | 1.33 1.03 |
| Dorsal fin base length | 55.4 13 | 54.3 | 51.5 | 56.6 | 1.45 |
| Pelvic fin length Pectoral fin length | 21.6 13 17.7 13 | 22.0 18.1 | 21.2 17.1 | 23.4 19.4 | 0.75 0.73 |
| Pre-dorsal distance | 33.4 13 | 32.4 | 31.1 | 34.7 | 0.98 |
| Pre-pectoral distance Pre-pelvic distance | 33.4 13 36.3 13 | 33.1 36.9 | 30.2 33.8 | 34.9 38.4 | 1.24 1.29 |
| Percent of head length Lower jaw length | 46.6 13 | 53.6 | 46.6 | 56.5 | 3.27 |
| Eye diameter | 18.7 13 | 24.0 | 18.7 | 28.3 | 2.24 |
| Snout length Interorbital width | 39.7 13 24.5 13 | 36.2 20.5 | 31.6 17.4 | 39.7 24.5 | 2.17 1.96 |
| COUNTS |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.