
Woolastookia basilicalabrica, Gerecke, 2014
|
publication ID |
https://doi.org/10.11646/zootaxa.3841.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:5FAADB3D-2359-453F-8BF5-1C2A33C6D178 |
|
DOI |
https://doi.org/10.5281/zenodo.6144817 |
|
persistent identifier |
https://treatment.plazi.org/id/03AD87B0-FF96-144E-FF46-BEFFFBA08056 |
|
treatment provided by |
Plazi |
|
scientific name |
Woolastookia basilicalabrica |
| status |
sp. nov. |
Woolastookia basilicalabrica sp. nov.
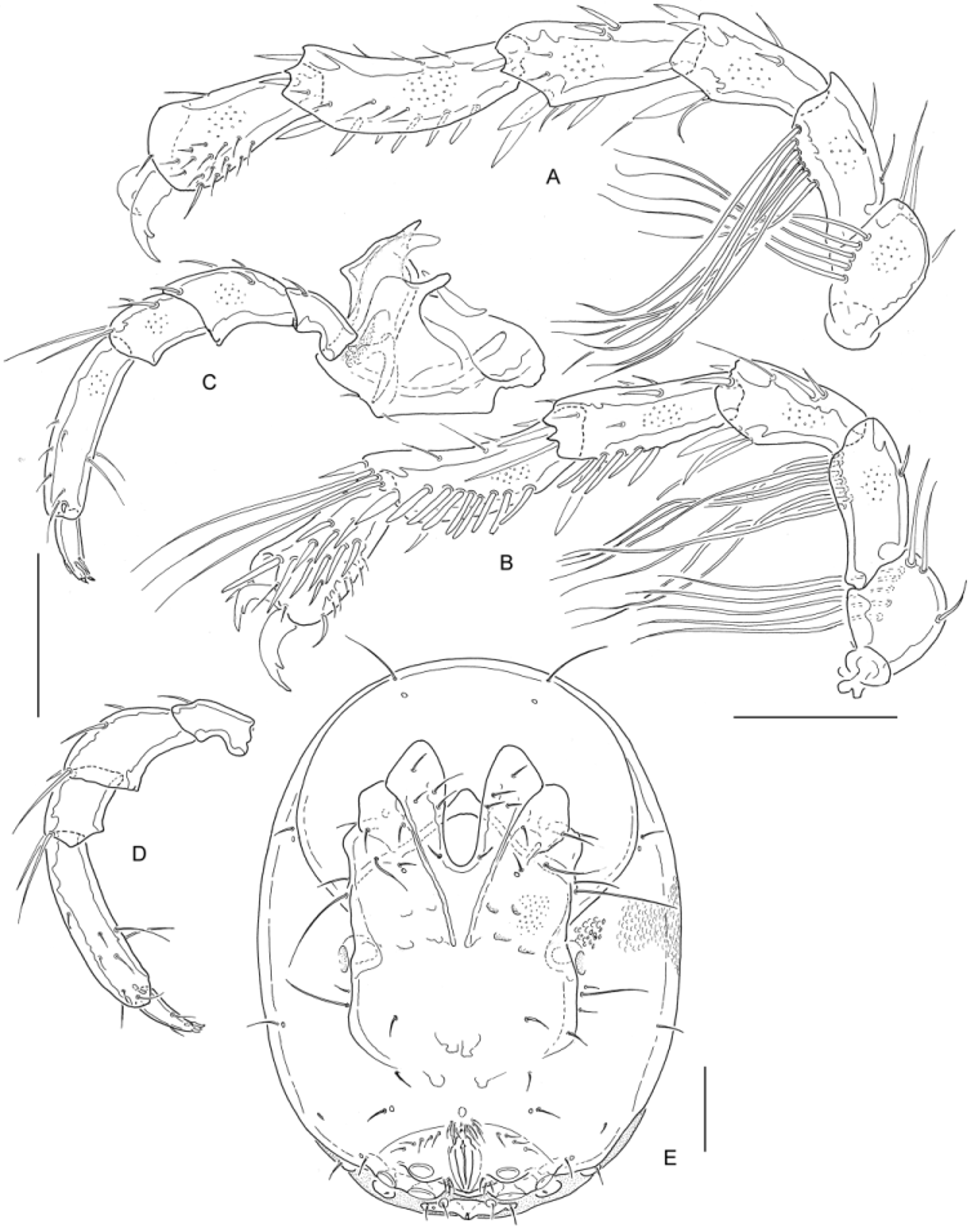
Fig. 27 View FIGURE 27
Material examined. Type series: Holotype ♂ NHMW, I 1115 b, Italy, Calabria, Pellegrino (CS), Saracena . Fonte Cannello, spring brook, 1385 m, WE 902 0 40, 0 8.10.1990 Gerecke. Paratypes, same site and date (2/7/0); I 1098, Basilicata Pollino (PZ), Terranova, spring N Lago Fondo, 1450 m, XE 0 52 228, 0 1.10.1990 (2/0/0).
Further material. I 106 Italy, Calabria, Sersale (CZ), Torrente Campanaro, 500 m, 19.07.1985 (0/2/0).
Diagnosis. Large in dimensions (idiosoma L ♂♂> 650, ♀♀> 620; palp total L ♂♂> 300, ♀♀> 280), idiosoma with slightly rounded lateral margins. Males with long, slender setae arranged in rows on IV-L-1/2.
Description. Colour brownish, sclerotized parts light violet. Idiosoma elliptical in outline, lateral margins rounded. Coxal field with parallel lateral margins extending from the III-L-insertion to halfway between IV-L insertion and genital field. One pair of setae free in the membrane posterior to genital field, excretory pore on separate plate bearing also a pair of flanking glandularia. Leg claws of I-III-L with slender blade and short, pointed dorsal and ventral clawlets, IV-L-claws with sexual dimorphism. Swimming setae numbers: II-L-4, 1; II-L-5, 3; III- L-4, 2; III-L-5, 3; IV-L-5, 3. Palp slender, similar in proportions in both sexes (relative L [% total L] P-1, 15–17, P- 2, 22–23, P-3, 12–14, P-4, 34–35, P-5, 14–15, L/H ratio P-1, 1.8–1.9, P-2, 1.5–1.6, P-3, 1.2–1.3, P-4, 3.6–4.1, P-5, 2.8–3.3; L ratio P-2/4, 0.63–0.66).
Male: IV-L-1 with 5 long, flagellar setae arranged in a line at anterior distal margin; IV-L-2 with a line of 6–7 similar setae in anterior half of ventral margin. IV-L-3 with equally diverging dorsal and ventral margins, distally forming sheaths (anteriorly triangular, with a short peg seta, posteriorly more rounded) covering the IV-L-4 base, distoventrally three setae, one strong, 2 more slender and longer; IV-L- 4 in shape similar to the preceding segment, but with more numerous strong distal setae extending over the posterior margin; IV-L-5 dorsal margin concave, ventral margin convex, on posteroventral margin a line of 8–10 heavy and stout setae, distally 3 swimming setae at posterior margin, and further strong, short setae; IV-L-6 stout, in distal part strongly enlarged, distal anterior side and distoventral margin covered by numerous fine setae, posterior surface with 10–12 strong, irregularly arranged setae, one of them in terminal position at the dorsal claw furrow margin; claw very strong, equally narrowed from the inflated basal part to the tip, without claw blade and dorsal clawlet, ventral clawlet short and stout. Genital field with setae in two groups; about 10 pairs along the area of fusion to ventral shield, 3 pairs flanking the posterior part of gonopore. Females: IV-L much more slender than in males, especially IV-L-4–6, except for the swimming setae on IV-L-5 without long setae, distal margins of IV-L-3/4 forming sheaths as in males; IV-L-2 with about 6 strong distoventral setae; IV-L-3 with several distal setae, one of them very strong, in distoventral position; IV-L-4 with a line of 5 strong setae on the distal ventral margin and several strong, long, distal setae (one of them swimming-setaline, on posterior distal margin); IV-L-5 only slightly curved dorsally, with a line of stronger setae in distal part of ventral margin and several very strong distal setae; IV-L-6 with little diverging dorsal and ventral margins, in distal half anteriorly and ventrally with a group of fine, hair-like setae, posteriorly several stronger setae in irregular arrangement; claws as on I–III-L.
Measurements: Male ( I 1115 b, 1089, n = 3): Idiosoma L/W 671–716/500–513, ratio 1.31–1.43; distance tip Cx-I – gonopore anterior end 464–468 (= 65–70 % idiosoma L). L/H IV-L-5, 132/43 (ratio 3.1), IV-L-6, 86/54 (ratio 1.6). Gnathosoma vL 102–110, chelicera L 124–131 (ratio L/H 2.7–2.8, basal segment/claw 1.7–1.9). Palp total L 319–323; L/H P-1, 52–56/24–28; P-2, 71–72/45–47; P-3, 40/35; P-4, 109–112/26–29; P-5, 44–45/14–15. Female ( I 1115 b, n = 3, mouth parts n = 1): Idiosoma L/W 630–711/450–513, ratio 1.31–1.50; distance tip Cx-I – gonopore anterior end 450–518 (= 71–73 % idiosoma L). Gnathosoma vL 107, chelicera L 135 (ratio L/H 2.9, basal segment/claw 1.8). Palp total L 286; L/H P-1, 44/42; P-2, 62/38; P-3, 39/31; P-4, 99/28; P-5, 43/15.
Discussion. Males of W. basilicalabrica differ from the two other species known from Europe, W. rotundifrons and W. minuta Pešić, Gerecke & Smit, 2010 , in the presence of groups of long and slender setae on IV-L-1/2. By their shape they remember "swimming" setae as they are described in many aturid taxa. However, typical swimming setae are in general found in both sexes, and also from their position close to the idiosoma, and the sexual dimorphism in claw shape, we should expect a different function, probably in mating and spermatophore transfer. A strong sexual dimorphism in length of setae, however inserting laterally on Cx-III, has been described for a species from China, W. foliatus Yi & Jin, 2012 . From most other points of view, there is a strong agreement with W. rotundifrons . Both sexes of the latter differ generally in minor dimensions (idiosoma L/W ♂♂ 500–585/ 383–441, ♀♀ 530–630/390–470, palp total L ♂♂ 230–290, ♀♀ 230–277), in single cases with a slight overlap in females. As this difference in size is the only possibility to distinguish female W. basilicalabrica from W. rotundifrons (e.g., there is a strong agreement in proportions of palp segments), the attribution of the two specimens from I 106 is uncertain. From our general experience with geographic variability of measurements characters in aturids, the detection of intermediate populations is probable in future.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |