
Paeonia lactiflora, Pall.
|
publication ID |
https://doi.org/10.1016/j.phytochem.2020.112573 |
|
DOI |
https://doi.org/10.5281/zenodo.8301284 |
|
persistent identifier |
https://treatment.plazi.org/id/03AD87AF-FFF8-FFB5-120F-FDB23C39FA3A |
|
treatment provided by |
Felipe |
|
scientific name |
Paeonia lactiflora |
| status |
|
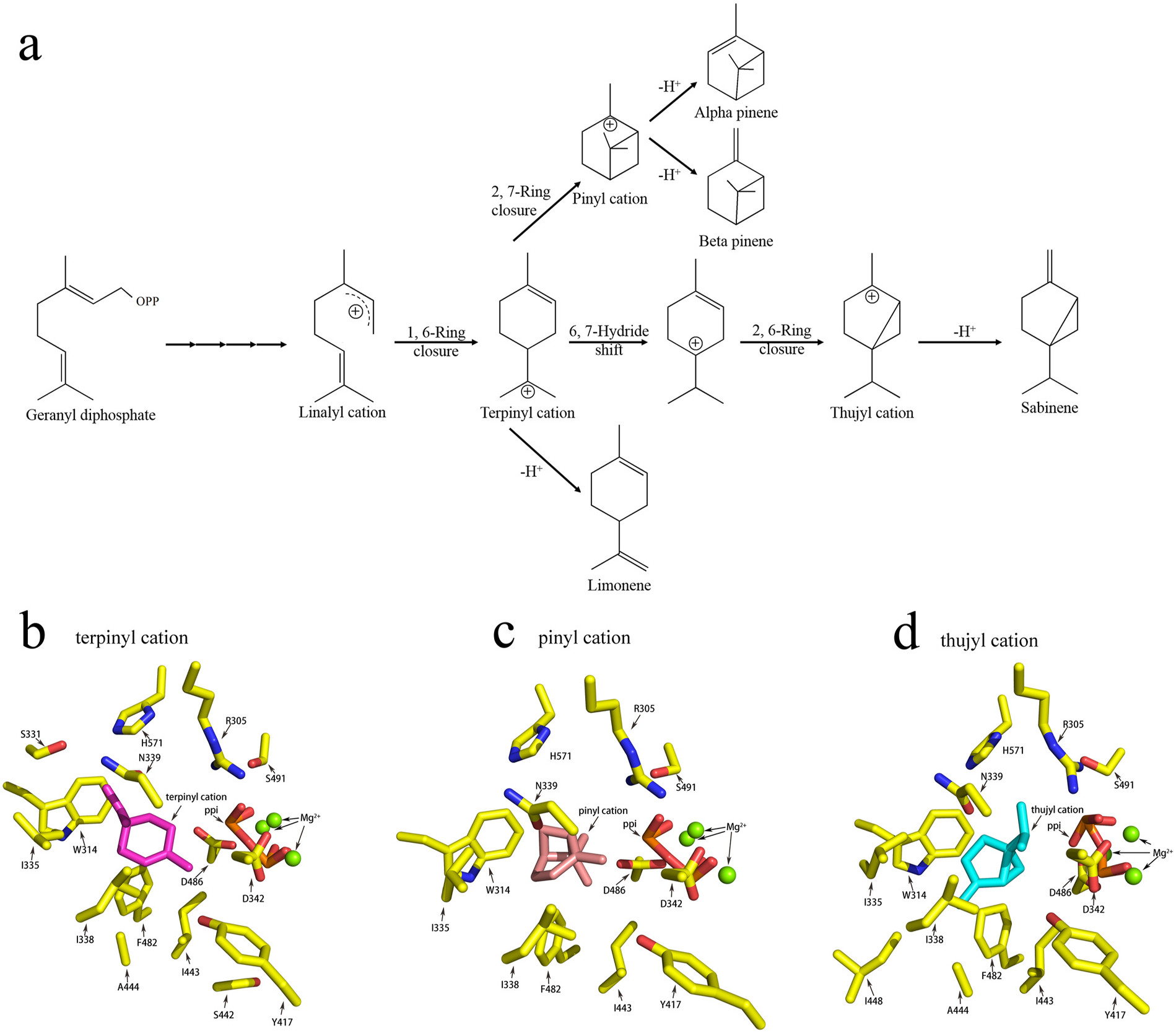
2.1. Homology modeling of P. lactiflora View in CoL View at ENA pinene synthase
The crystal structures of pinene synthases have not been reported. Therefore, we constructed a homolog model of P. lactiflora pinene synthase using the crystal structure of S-limonene synthase as the template. In order to investigate the catalytic mechanism, we docked three carbocation intermediates (terpinyl, pinyl and thujyl cations) into its active site ( Fig. 1B–D View Fig ). The docking results suggest that carbocations bind to a similar location in the active pocket. Nearby residues include R305, S311, W314, I335, I338, N339, D342, Y417, I443, A444, I448, F482, D486, S491 and H571. Some of these residues could make hydrophobic interactions with the carbon skeleton of carbocations. Certain polar or aromatic residues may stabilize the positive charge and influence the product profile of the enzyme.
2.2. Alanine scanning mutagenesis identifies mutations that cause an increase in sabinene production
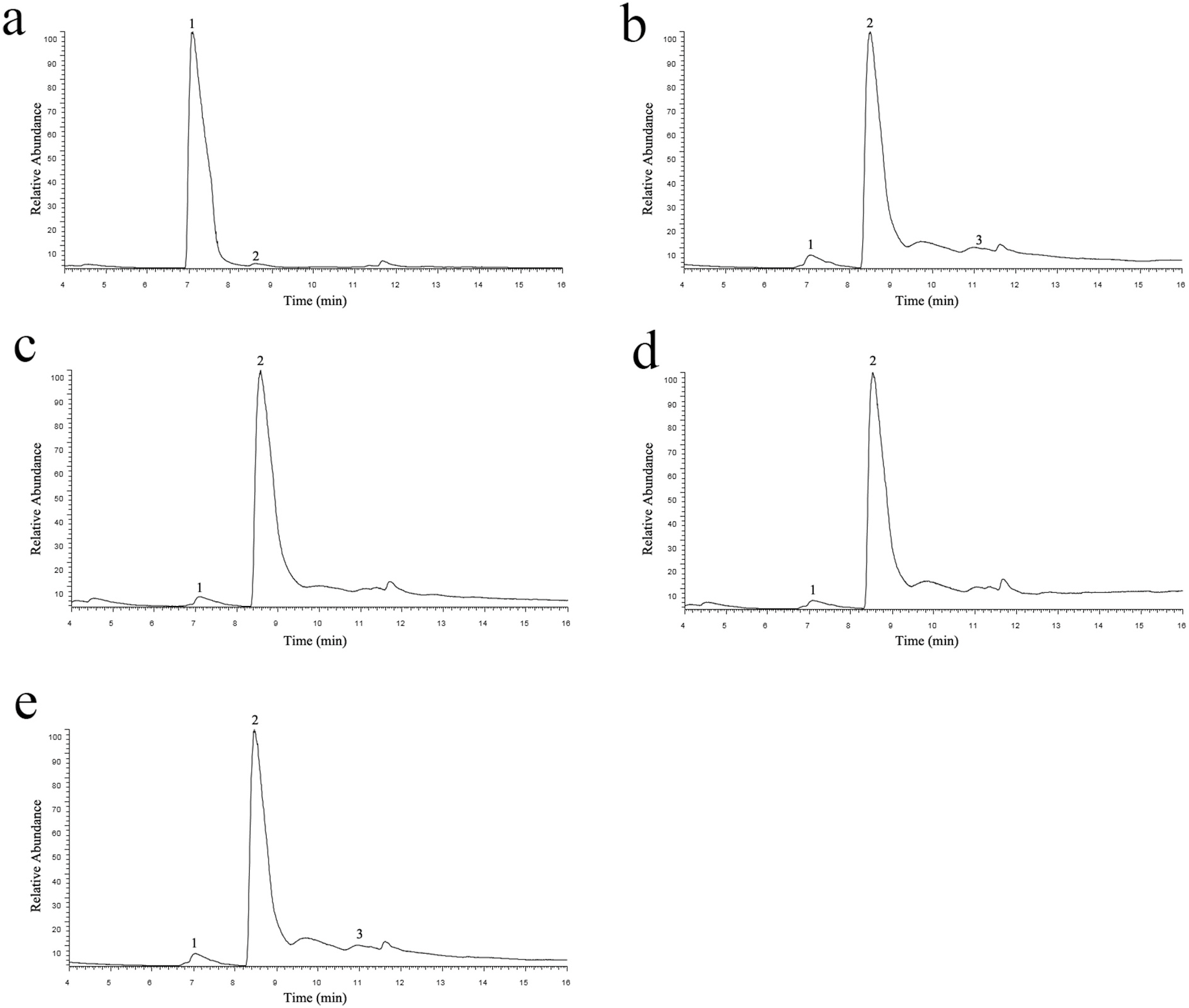
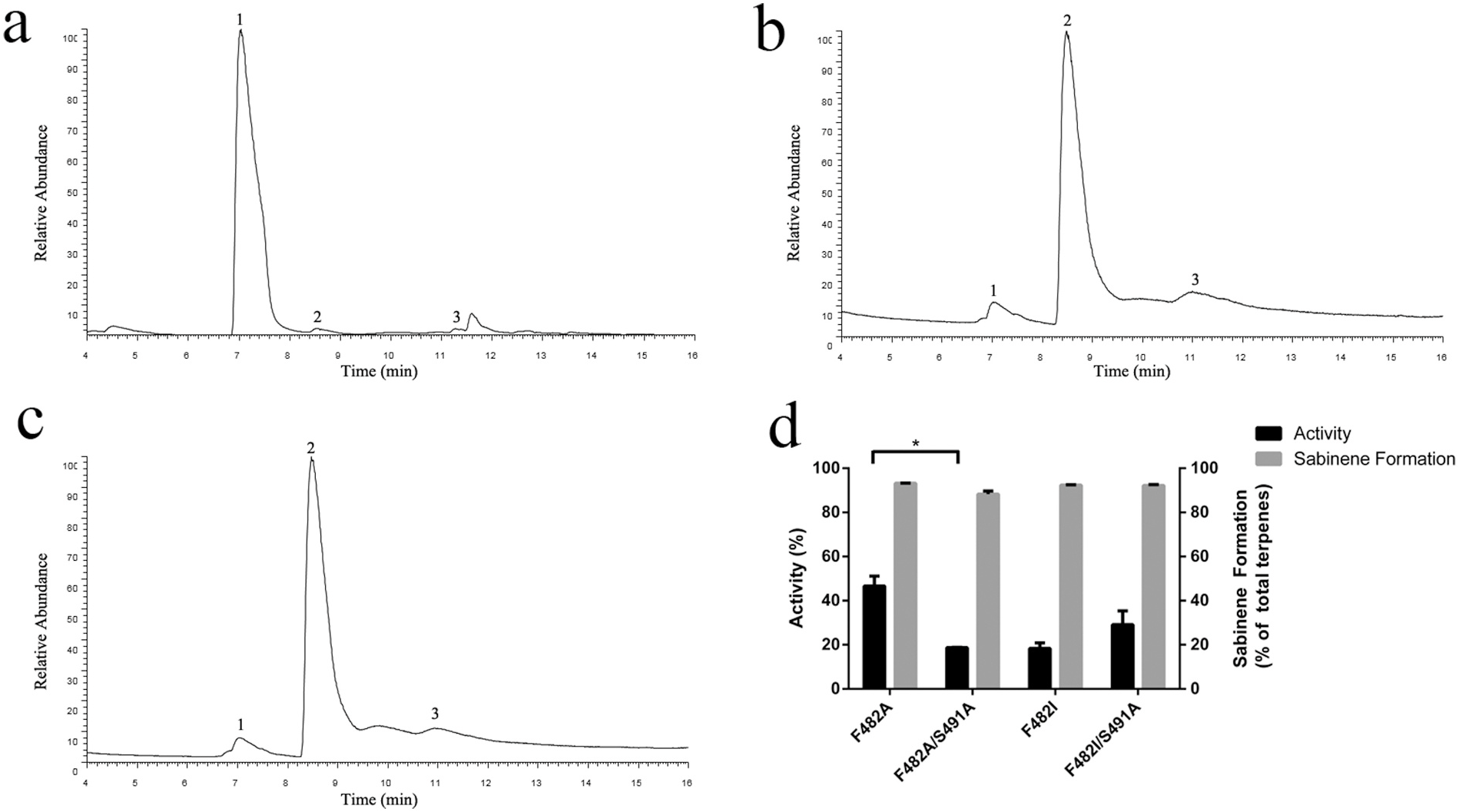
We first identified residues within 5 Å from the docked carbocations and performed alanine-scanning mutagenesis on them. Most of these mutations do not show dramatic changes in their product profiles. However, two mutations show interesting alteration in the product profile. One of them, F482A, essentially converts the enzyme to sabinene synthase. About 90.8% of its products is sabinene, while 97.7% of the products of the WT enzyme is α- pinene ( Table 1 View Table 1 , Fig. 2A–B View Fig ). Another mutation, I335A, also causes an increase in sabinene production, though not as dramatic as F482A ( Table 1 View Table 1 ). Importantly, although both mutants show an altered product profile, their overall activities are only moderately decreased ( Table 1 View Table 1 ).
2.3. Phenylalanine at the 482 position is required for pinene production
We hypothesized that the aromatic ring of F482 might play a role in stabilizing the pinyl cation. To investigate this possibility, we mutated F482 to Val and Ile, two nonpolar residues with different side chain sizes. We also mutated it to Thr, Arg and Trp, representing polar, charged or non-Phe aromatic residues. The results suggest that any mutations that changes the phenylalanine at this position leads to an increase in sabinene production and almost abolishes the α- pinene production ( Table 1 View Table 1 ). Replacing phenylalanine with nonpolar residues (F482I, F482V) all effectively convert the enzyme to sabinene synthase ( Fig. 2C–D View Fig ). More than 90% of products made by these mutants are sabinene ( Table 1 View Table 1 ). Interestingly, F482T also predominantly produces sabinene, which seems to suggest that a polar residue here is not sufficient to stabilize the pinyl cation ( Table 1 View Table 1 , Fig. 2E View Fig ). Other mutations (F482Y, F482R and F482W) retain very little activities ( Table 1 View Table 1 ).
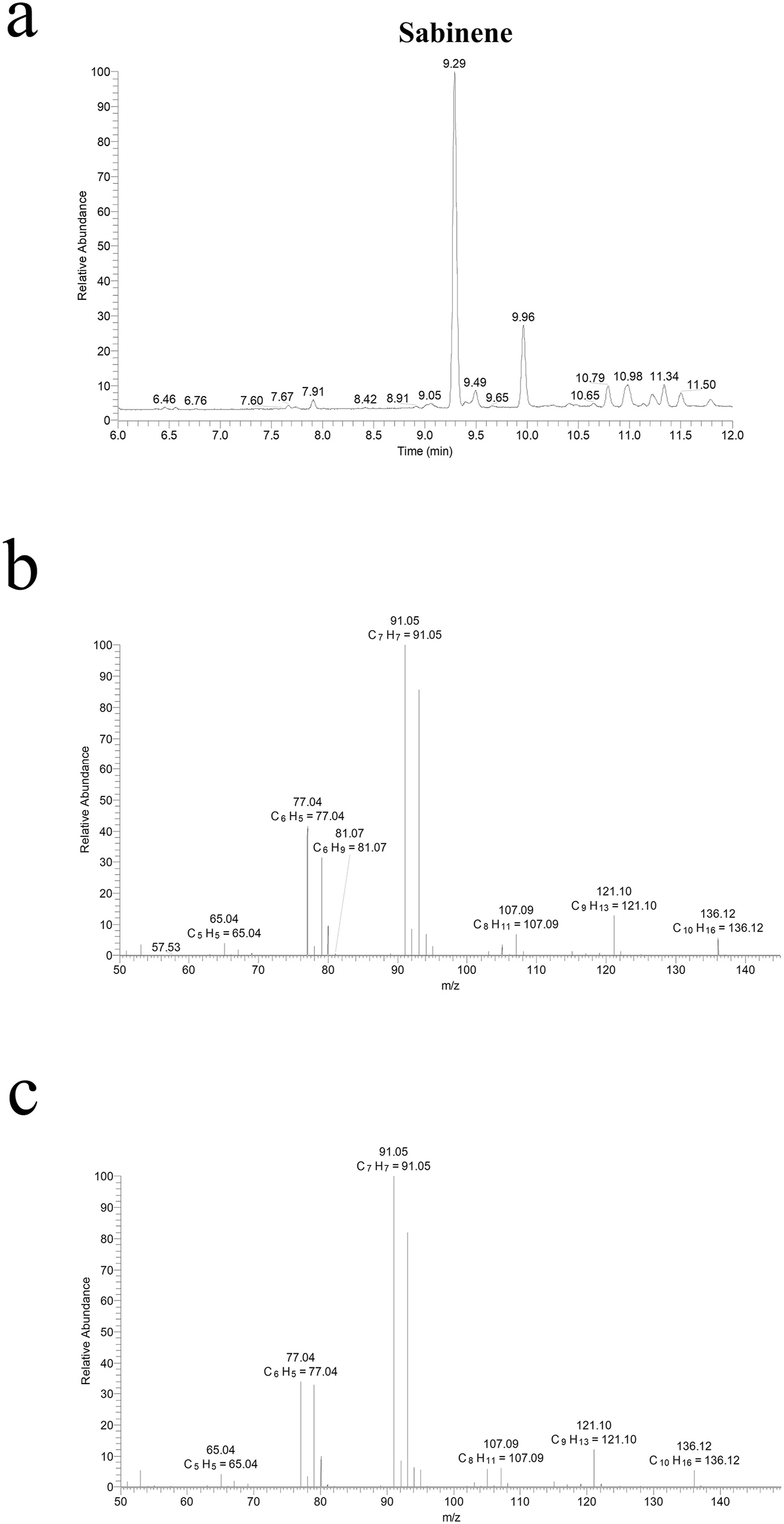
In order to verify that the mutant enzymes indeed produce sabinene, we analyzed the reaction product of F482L using the high resolution hybrid quadrupole-orbitrap GC-MS/MS system ( Fig. 3A–C View Fig ). By comparing with the sabinene standard, the peak eluted at 9.29 min was identified as sabinene. Its mass spectrum was measured and could be readily matched with that of sabinene standard ( Fig. 3B–C View Fig ).
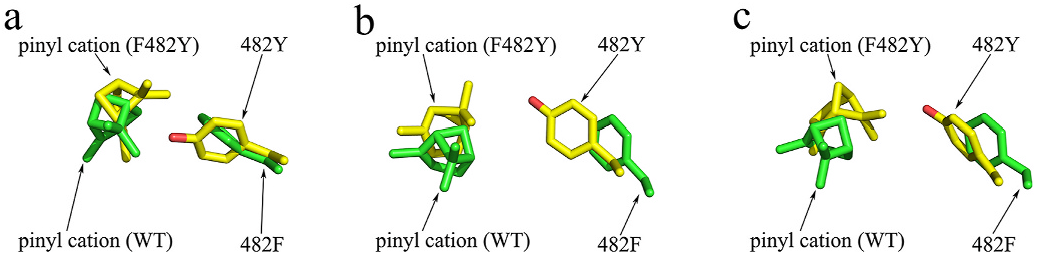
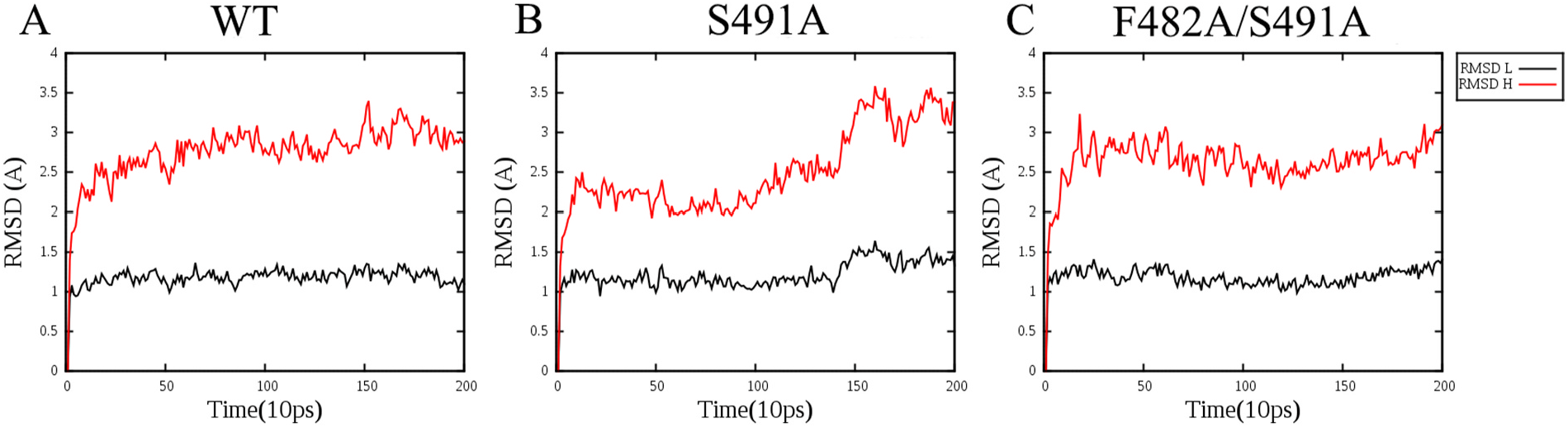
It is interesting to note that replacement of F482 to other aromatic residues (Tyr or Trp) causes a drastic decrease in enzyme activities. To investigate this issue, we first docked the pinyl cation into the active pockets of WT and F482Y. It could be seen that in these two structures, the conformation of the docked pinyl cation is quite different ( Fig. 4A View Fig ). We then performed a 2 ns molecular dynamics simulation. The results suggest more conformational changes ( Fig. 4B–C View Fig ). Therefore, the extra hydroxyl group in Tyr could significantly change the conformation of the carbocation, which may explain the decreased activity and altered product profile.
2.4. S491A mutation enhances the overall activity of pinene synthase without changing its product profile
During our Ala scanning experiments, we identified a mutation S491A that enhances overall activities ( Fig. 5A, D View Fig , Table 1 View Table 1 ). The overall activity of S491A mutant is about 29% higher than that of the WT enzyme ( Table 1 View Table 1 ). Furthermore, S491A does not change the product selectivity, as more than 90% of their products remains α- pinene ( Fig. 5A View Fig , Table 1 View Table 1 ).
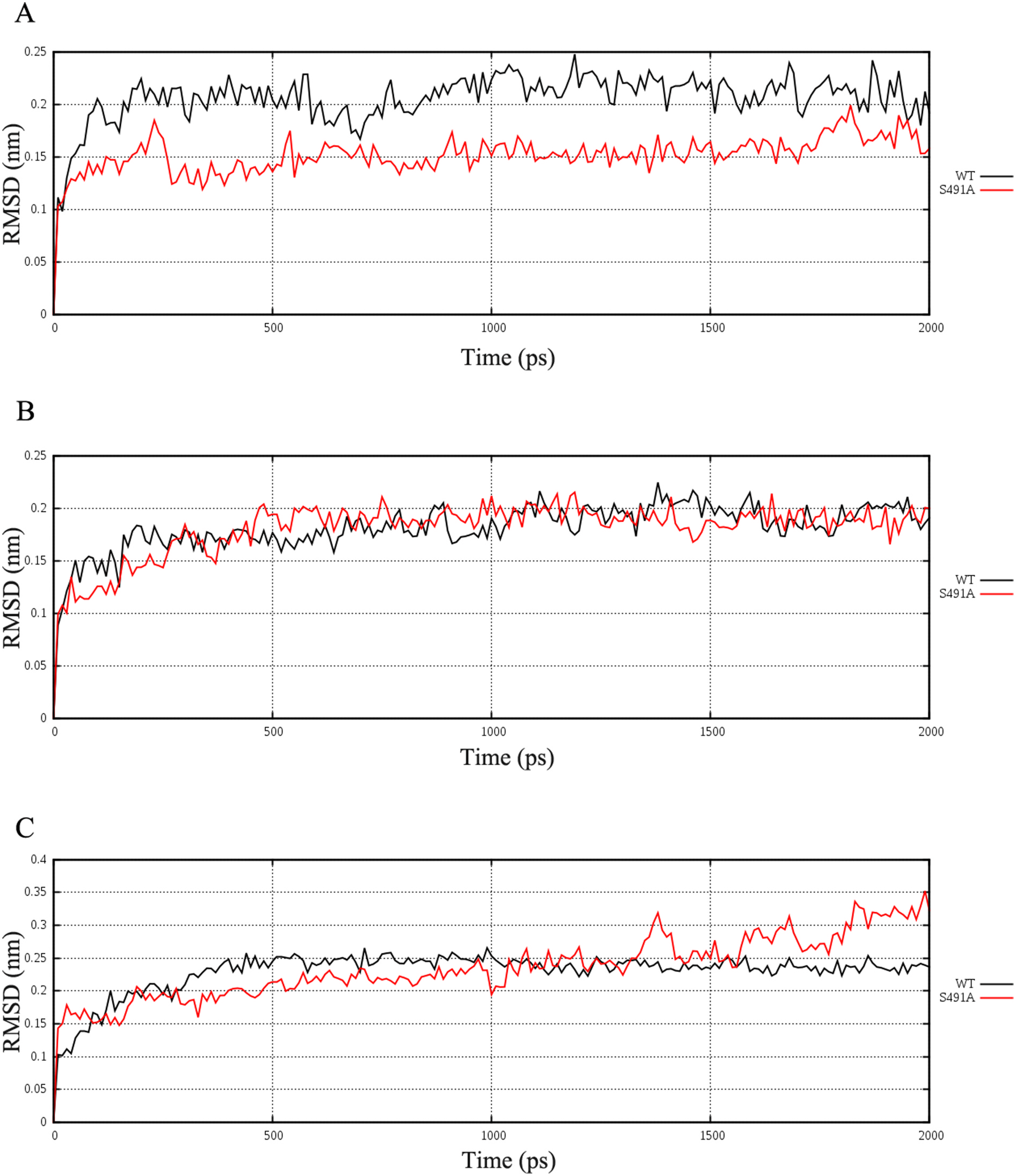
What is the mechanism of S491A to enhance the enzymatic activity? Since S491A still predominantly produces pinene, we docked the pinyl cation into the active pocket of the mutant enzyme and performed a 2 ns of molecular dynamics simulation. We also performed the same dynamic simulation on the WT enzyme in complex with the pinyl cation. Our data suggests that the active site residues in S491A have a much smaller RMSD than those in the WT enzyme, indicating the mutation S491A could cause the active pocket to be more rigid ( Fig. 6A View Fig , STable 2). Interestingly, simulation on the WT or S491A enzymes in complex with the terpinyl or thujyl cation does not show the same differences in RMSD values of active site residues ( Fig. 6B–C View Fig , STable 2). This probably indicates that the S491A mutation increases the rigidity of the active pocket only when it binds to the pinyl cation.
To better quantify the mobility of protein structures, we reanalyzed the simulation data using MDLovoFit ( Martínez, 2015). This way we could subdivide the protein into regions with high and low mobility ( Fig. 7A–C View Fig , STable 3). Interestingly, in S491A, fewer residues are classified as of high mobility than residues in WT enzymes (59% vs 63%). Furthermore, for residues classified with the highly mobile subset, those in S491A have lower average RMSD than those in WT proteins ( 0.253 nm vs. 0.278 nm). Therefore, there is a slight but significant increase in rigidity in S491A mutant protein.
2.5. Combination of S491A with F482 mutants has mixed results in the overall activity
Since all four sabinene synthase mutants show some decreases in the catalytic activity, we wondered whether we may restore their activities to that of the WT pinene synthase by combing with S491A. We produced double mutants F482A/S491A and F482I/S491A. Both seem to retain the same product selectivity of the original single mutants, though F482A/S491A shows an insignificant decrease in sabinene production (P =0.39; Fig. 5B–C View Fig , Table 1 View Table 1 ). In overall activities, F482I/S491A shows an insignificant increase in activity (P =0.27), while F482A/S491A has a significant decrease in activity (P < 0.05; Fig. 5D View Fig , Table 1 View Table 1 ). Therefore, it seems that the effect of combining two mutants is more complicated than we expected.
We asked whether the combination of F482A and S491A mutations could affect the rigidity of the protein. After a 2 ns of molecular dynamics simulation and data analysis using MDLovoFit, we found the rigidity of F482A/S491A is similar to that of S491A (STable 3). Apparently, the decrease of activity by the combination of F482A with S491A cannot be explained by the overall RMSD change.
To further investigate this issue, we analyze the RMSF values based on the simulation data (SFig. 1A–C). Compared with S491A and WT, F482A/S491A shows an RMSF spike at a region between residues 219 and 223. In the structural model, this region forms a loop in the protein structural model (SFig. 2). However, this loop is quite far from the active site (about 40 Å). Therefore, its mobility could only affect the enzyme activity indirectly. For example, it could influence the closure of the active pocket.
We also superimposed the structural model of WT, S491A and F482A/S491A in complex with the docked pinyl cation (SFig. 3). In the structural model, the pinyl cation is docked between the residue 491 and 482 (SFig. 3). Interestingly, in WT and S491A, the position of the pinyl cation is similar, while in F482A/S491A, the pinyl cation is shifted towards the A482. This change in the carbocation position may impair the enzyme activity (SFig. 3).
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |