
Balaenidae Gray, 1821
|
publication ID |
https://doi.org/ 10.24199/j.mmv.2005.62.2 |
|
DOI |
https://doi.org/10.5281/zenodo.10870916 |
|
persistent identifier |
https://treatment.plazi.org/id/03ACB35A-DC76-FF9C-FF7A-F913FEDEF904 |
|
treatment provided by |
Felipe |
|
scientific name |
Balaenidae Gray, 1821 |
| status |
|
Family Balaenidae Gray, 1821 View in CoL
Genus and species indeterminate
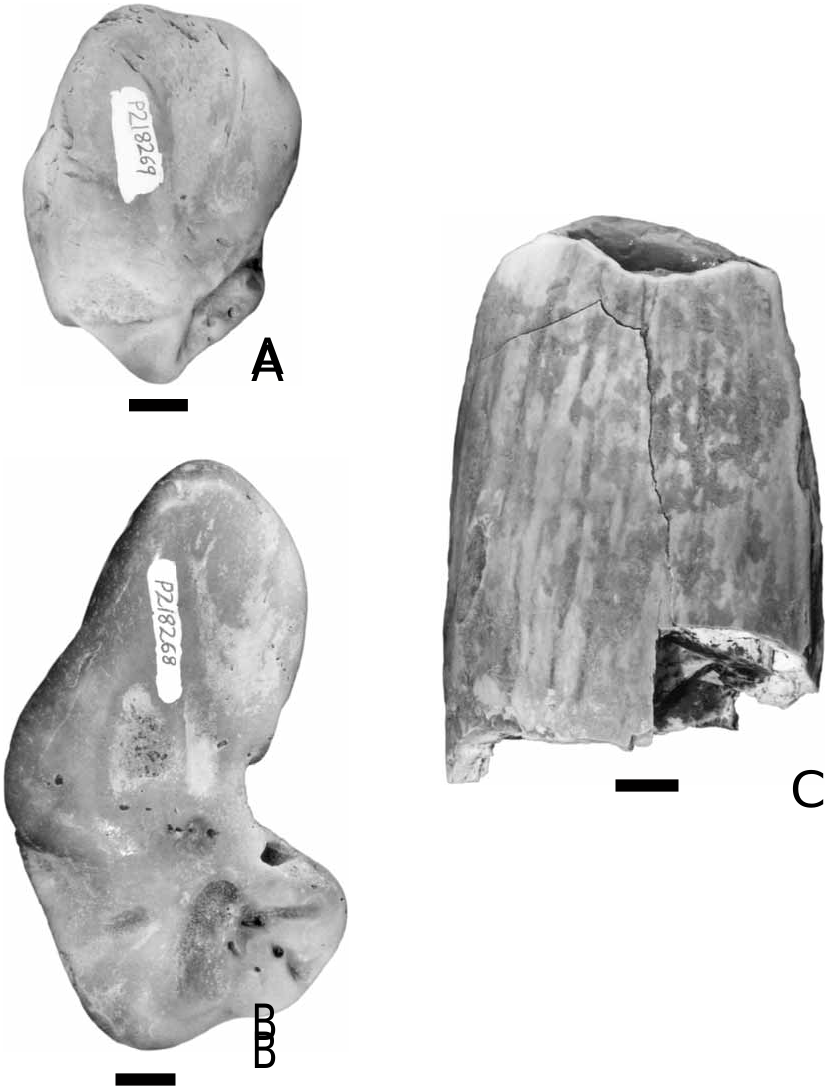
Referred specimen. NMV P218269, incomplete right periotic; anterior and superior processes virtually complete, but pars cochlearis almost entirely worn off, and only anteriormost base of posterior process preserved ( Fig. 3A View Figure 3 ).
Description. P218269 is highly polished and abraded. The anterior process is blunt and globose, being indistinct from the superior process. There is marked lateral exostosis of the superior process lateral to the epitympanic recess. The lateral aspect of the anterior process is rugose and pitted. Posteriorly, this pitting decreases in density. Only the lateralmost region of the pars cochlearis is preserved. In medial view, the most notable feature is the sulcus for the facial nerve (cr. VII), the course of cr. VII being preserved from its entry into the body of the periotic at the aperure of the internal facial foramen, to its ventral exit into the epitympanic cavity via the ventral facial foramen. All other features of the pars cochlearis and epitympanic recess have been obliterated. Posterior to the broad and shallow hiatus epitympanicus is a remnant of the base of the posterior process (which is directed posterolaterally and somewhat ventrally).
Discussion. Miller (1924: 8–9) listed the following features that distinguish the periotics of Balaenidae from those of Balaenopteridae (and other baleen-bearing Mysticeti ): (1) axis of anterior process of periotic parallel with axis of internal acoustic meatus; (2) [longitudinal] axes of anterior and posterior processes converge at an acute angle; and (3) pars cochlearis small relative to rest of periotic. In addition to the preceding features, the possession of massive lateral exostosis of the anterior process and anterolateral superior process, such that the anterior process appears swollen (as noted by Fordyce, 1982: 48), seems to be a feature shared by all extant and late Neogene balaenid periotics. It is largely on the basis of the latter character and the phenetic similarity of P218269 to a periotic (P16195) from the Lower Pliocene Black Rock Sandstone of Beaumaris identified as belonging to cf. “ Balaena ” ( Gill, 1957) that P218269 is referred to Balaenidae , genus and species indeterminate.
The fossil record of Balaenidae begins in the Late Oligocene (c. 28 Ma: Fordyce, 2002b), although the record only becomes reasonably well known from the Mio-Pliocene boundary onwards ( McLeod et al., 1993; Bisconti, 2003). Morenocetus parvus Cabrera, 1926 is the geologically oldest named balaenid, from the early Early Miocene (Aquitanian) of Patagonia. From the end Aquitanian to early Tortonian of the Miocene the evolutionary history of Balaenidae is virtually unknown. The extant balaenids include Balaena mysticetus Linnaeus, 1758 , Eubalaena australis Desmoulins, 1822 , E. glacialis Müller, 1776 , and E. japonica Lacépède, 1818 (e.g., Cummings, 1985; Reeves and Leatherwood, 1985; Bannister, 2002). Note that Rice (1998) included all extant balaenids in the genus Balaena and recognised only two species, B. mysticetus and B. glacialis . The taxonomic scheme of Bannister (2002) is used herein. Balaena is known from the Early Pliocene of the North Atlantic ( McLeod et al., 1993; Westgate and Whitmore, 2002). There are very few confirmed pre-Quaternary fossil records of Eubalaena . Bisconti (2003, 2005) referred the Pliocene Balaena belgica Abel, 1941 to Eubalaena belgica . McLeod and others (1993: 63) suggested that a balaenid periotic from the Early Pliocene of South Australia (originally recorded by Howchin: 1919) could represent Eubalaena (as opposed to its original referral to Balaena ). Dixon (1990) described an incomplete Recent Eubalaena australis skeleton from Altona Bay, near Melbourne, Victoria. The latter specimen (C27879) includes tympanics and periotics. The extinct genera Balaenula and Balaenotus have been recorded from the Late Miocene through Pliocene of the N Pacific ( Barnes, 1977; McLeod et al., 1993) and N Atlantic ( McLeod et al., 1993; Bisconti, 2003 and references therein). Recently, Bisconti (2005) described a new genus and species of relatively small balaenid, Balaenella brachyrhynus , from the Early Pliocene of Belgium.
The incompleteness of P218269 and lack of information on the extent of intraspecific and ontogenetic variation in balaenid periotics, hampers comparisons with described extant and fossil balaenid taxa. Furthermore, there are as yet no published criteria for discriminating between the periotics of Balaena and Eubalaena . Despite these problems, it may be noted that P218269 is similar in overall size to several isolated balaenid periotics from the uppermost Miocene to Lower Pliocene Black Rock Sandstone and Grange Burn Formation of Victoria (e.g. P16195, P48865, P160438, and P197824). The discovery of a more complete periotic (including the pars cochlearis) is necessary before any further comparisons between the Portland Pliocene balaenid and the other Victorian specimens listed above can be made.
| NMV |
Museum Victoria |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
SubOrder |
Mysticeti |
|
Family |