
Liolaemus hermannunezi, Pincheira-Donoso, Daniel, Scolaro, Alejandro & Schulte, James A., 2007
|
publication ID |
https://doi.org/ 10.5281/zenodo.176292 |
|
DOI |
https://doi.org/10.5281/zenodo.6249795 |
|
persistent identifier |
https://treatment.plazi.org/id/03AB87EA-243A-7531-45A9-425286C7F83A |
|
treatment provided by |
Plazi |
|
scientific name |
Liolaemus hermannunezi |
| status |
sp. nov. |
Liolaemus hermannunezi sp. nov.
Figures 2 View FIGURE 2 and 4 View FIGURE 4
Liolaemus rothi Donoso-Barros (1974:287) ; Núñez and Jaksic (1992:76); Pincheira-Donoso (2003a:18); Schulte et al. (2004:410).
Liolaemus (Eulaemus) rothi Pincheira-Donoso and Núñez (2005:99) .
Type material. Holotype. MNHNC-3785, male, collected 10 km E from Los Barros (37°31’S; 71°15’W) on the road to Pichachén Pass, Eighth Administrative Region, Chile, 18 November 2001 by D. Pincheira- Donoso.
Paratypes. MNHNC-3501, male, same locality as holotype, collected 10 February 2001 by J. A. Schulte II and J. P. Valladares; MNHNC-3502, 3503, both females, same data as male paratype; JAS-DC-718, female, same data as holotype; CHDPD-995, female, same data as holotype.
Etymology. This species is dedicated to Herman Núñez, from the National Museum of Natural History of Chile, in recognition of his great contribution to the systematics and ecology of Chilean lizards of the genus Liolaemus . DP-D particularly thanks his teachings, his intimate friendship and his persistent help and encouragement.
Diagnosis. A small bodied Liolaemus species belonging to the boulengeri clade diagnosed by Etheridge (1995), as having a femoral patch of abruptly enlarged scales on the posterior surface of the thigh, and by the hypertrophied puboischiotibialis muscle (recently recognized as flexor tibialis internus; Abdala et al. 2006), a trait well-developed in adult males. Into the boulengeri clade, Etheridge (1995, 2000) also recognized the wiegmannii species group, which is characterized by lorilabials distinctly smaller than supralabials, usually in two rows between the subocular and supralabials, with the posteriormost not elongate, with sublabials contacting the mental shield, which is widest posteriorly, and infralabials flat to concave. However, traits of the wiegmannii group are not observed in L. hermannunezi , and morphology and coloration differentiate L. hermannunezi from most of the remaining species belonging to the L. boulengeri clade, except L. loboi , L. rothi , L. sagei and L. tehuelche . Liolaemus hermannunezi differs from L. tehuelche in having a black belly, a smaller body size (49.1–64.1 mm; mean = 53.1 mm), and a distribution restricted to the Laguna del Laja in Chile (37°31’S; 71°15’W), whereas L. tehuelche is only black on the throat, has a larger body size (55.5–74.2 mm; mean = 66.4 mm), and a more austral distribution in Argentina (40°44’S; 70°34’W). Liolaemus hermannunezi differs from L. sagei in having a smaller body size (see above), larger and fewer body scales (Table 1), and has ventral melanism, mainly in abdominal and gular areas, whereas L. sagei is uniformly grey or blue ventrally. From L. rothi , L. hermannunezi differs in having a smaller body size (SVL over 100 mm in L. rothi ; see above), smaller and therefore more body scales (Table 1), and a black belly in males, restricted to the gular region in females. This last trait is absent in L. rothi (Table 1). Finally, L. hermannunezi differs from L. loboi in having a smaller body size (see results for statistical comparisons), a higher number of scales around the middle of body (range in L. hermannunezi = 66–77, mean = 71.8; versus range in L. loboi = 59–70, mean = 63.7), is black ventrally, and has a more boreal distribution in Chile, whereas in L. loboi ventral melanism is conspicuous, and occurs in more austral areas in Argentina (see Results for details, also Fig. 5 View FIGURE 5 , Table 1).
Description of the Holotype – A small lizard with a relatively stout body. Snout-vent length 53.3 mm, axilla-groin distance 24.0 mm. Head slightly longer (12.6 mm) than wide (10.7 mm). Head height 7.8 mm. Snout moderately large. Neck as wide as head, with a shallow lateral fold, extending from posterior edge of external auditory meatus to insertion of the forelimb, forming an antehumeral fold and deep antehumeral pocket. Dorso-cervical and hemigular folds absent. Forelimb length 22.2 mm. Hind limb length 34.9 mm. Toe tips of adpressed hind limb reach posterior edge of external auditory meatus. Tail 88.7 mm long, cylindrical, longer than snout-vent length.
Dorsal head scales slightly convex and smooth. Rostral pentagonal, expanded laterally, contacting six smaller scales. Two tiny pentagonal postrostrals. Nasal contacting seven smaller scales, surface twice as large as interparietal shield. A single row of four azygous scales surrounded by 10 frontonasals. Two prefrontals. Frontal divided transversely and longitudinally into seven small scales of similar size, 3–4 times smaller than prefrontals. Three postfrontals arranged in a transverse row, directly in contact with interparietal, hexagonal, and slightly larger than postfrontals. Parietals irregular, twice as large as interparietal, and with a smaller scale inserted between them. Occipital scales homogeneous, small, irregular, juxtaposed. Supraorbital semicircles convex. Circum orbitalis consisting of 13–13 small scales. Supraoculars 6–6, hexagonal, and wider than long, larger than other dorsal head scales. Superciliaries imbricate. Two rows of scales between supraoculars and superciliaries. Upper-ciliary scales 15–14, and 12–10 lower-ciliaries. Suboculars and postoculars enlarged, first one contacting an irregular polygonal preocular. Loreal region slightly concave, covered by three small scales, medial larger than other two. Supralabials 7–7, posteriors three times longer than anteriors. A single and irregular row of loreolabial scales between subocular and supralabials. Temporal subcircular, smooth, and subimbricate. Frontonasal, loreals, canthal, supralabials, loreolabials, temporals, and supraorbitals with tiny scale organs.
Mental subtriangular, slightly wider than rostral, bordered by two lower labials and two postmentals. Two divergent rows of 7–8 postmental scales, posteriors smaller than anteriors. Infralabials 6–7, larger than supralabials, smaller than those in posterior region. Single row of sublabials between postmentals and infralabials, that turn into two rows under second infralabial and in three rows under fifth infralabial. Gulars subcircular, smooth, and imbricate. External auditory meatus higher than wide, smaller than orbit, surrounded by several tiny granular scales, with two large scales along anterior margin, and conspicuous auricular scale in dorsal margin.
Dorsal body scales subtriangular and subimbricate, strongly keeled, never mucronate, lacking interstitial granules. Scales of body flanks similar to dorsals, but become smooth, with tiny interstitial granules. Ventral body scales 1.5 times larger than dorsals, rhomboidal, smooth, and imbricate, subtriangular and smaller in precloacal region. Scales around midbody 66. Lateral neck folds covered by several tiny granular scales. Similar scales on shoulders, axilla, and cloacal regions.
Forelimb scales subtriangular, imbricate, and slightly keeled, becoming smooth on dorsal surface of front feet. Lower humeral region covered by granular, juxtaposed tiny scales, becoming larger and subimbricate on forearm. Hind limbs with rhomboidal, slightly keeled, and subimbricate-imbricate scales, with a few tiny interstitial granules. Ventral scales of hind limbs rhomboidal, smooth, and imbricate. Coxal scales never projected. Posterior surface of thighs with a patch of abruptly enlarged scales. Twenty six strongly keeled subdigital lamellae on the fourth toe of right foot. Dorsal and lateral caudal scales rhomboidal, strongly keeled, becoming distinctly rectangular on distal tail. Ventral scales of the tail subtriangular, smooth, and imbricate, longer and keeled distally. Nine precloacal glands.
Coloration (on the basis of the entire type series) – The ground colour in living specimens is grayish or brownish olive on the dorsal head and body surfaces, becoming gray or blue in preserved animals. Head with irregular and small brown, black, and white spots. Suboculars and supralabial scales white, with indistinct gray spots. Two fine (1–2 scales wide) white dorsolateral stripes run from superior margin of nasals, to tail base. Nine subcircular paravertebral black spots, similar or larger in size than orbit (each 19–25 dorsal scales), bordered by gray and white tiny marks, and arranged alternately or in pairs, in two parallel and longitudinal rows, most conspicuous at tail base. Prescapular and postcapular spots black and conspicuous. Fine (1–2 scales wide) white stripe on dorsal neck, normally fragmented on the back. Flanks with white ventrolateral stripe and black spots, slightly smaller than paravertebral ones, and bordered by tiny white and blue irregular spots in upper region. Flanks with distinct yellow scales in life. The same pattern is represented on the dorsal tail surface, being clearly more irregular. Distinct black, grey, and white spots are irregularly distributed on the dorsal surface of limbs.
Pale gray-whitish ground colour ventrally, but black on mid-belly and gular surface. Lower ventrolateral field irregularly scattered with small brown marks. Ground colour of ventral limbs similar to ventral body. Ventral tail unspotted, with small and indistinct gray marks in postcloacal region. Precloacal region pale. Precloacal glands orange. In preserved specimens ventral colour fades pale blue.
Variation. No obvious differences were detected between sexes, except in precloacal glands, developed in males and totally absent in females. Also, ventral and gular melanism is slightly more conspicuous in males.
Frontonasal azygous 2–4, frequently with small supernumerary scales. Frontal region transversely and longitudinally divided, resulting 6–8 similarly sized small frontals. Single transverse row of 3–5 postfrontals behind frontal. Parietal scales irregular, smaller, equal, or larger than interparietal shield, frequently separated by tiny supernumerary scales. Supraorbital semicircles surrounded by 13–16 small scales. Supraoculars 4–7 on each supraorbital semicircle. Upper ciliaries 13–15, and 10–13 lower ciliaries. Loreals 3–6, polygonal, variable in size. Anterior supralabials similar or larger than the posterior ones, 7–9 on each side; 2–3 enlarged scales on anterior margin of external auditory meatus. Single auricular shield differentiated. Two divergent rows of 5–8 postmental scales contacting mental. Infralabials 6–7, similar or slightly larger than supralabials.
Dorsal body scales rhomboidal or subtriangular, keeled, subimbricate or juxtaposed, often with interstitial granules. Scales around midbody 66–77. Ventral body scales with negligible variation. Dorsal scales of forelimbs rhomboidal or sublanceolate. On hind limbs, dorsal scales rhomboidal, slightly keeled, imbricate or subimbricate, with interstitial granules. Fourth toes with 24–27 subdigital lamellae. Dorsal tail scales subcircular or subtriangular, keeled and imbricate, becoming more polygonal medially. Ventral scales of tail subcircular or subtriangular, smooth and imbricate, becoming more triangular and keeled medially. Precloacal glands 7–9, restricted to males.
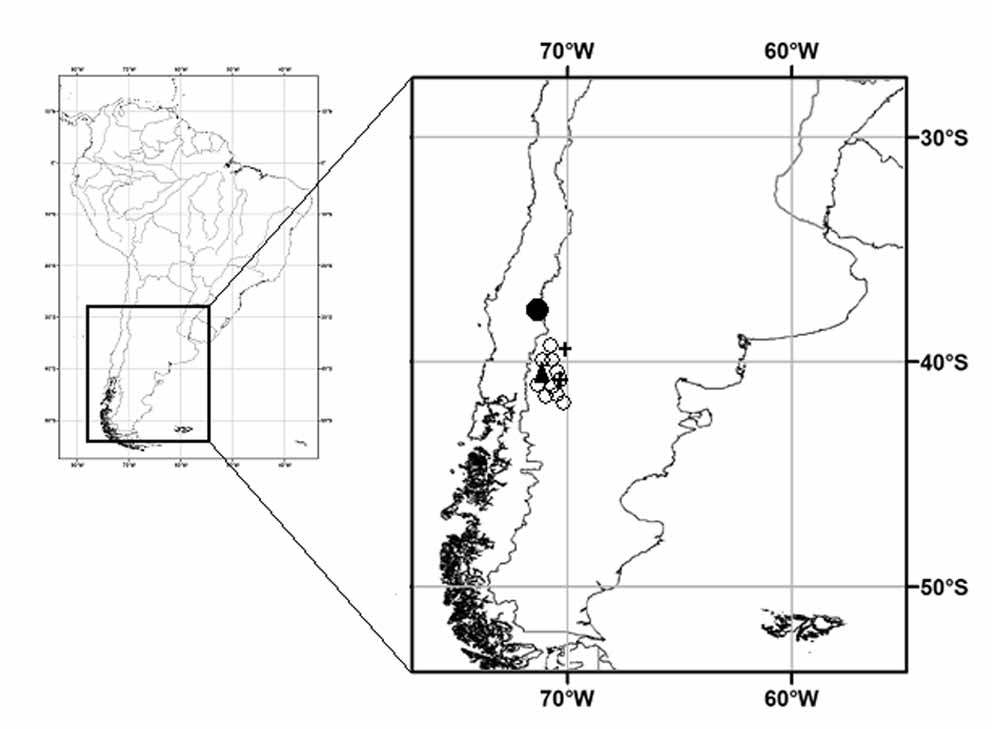
Geographic distribution. Liolaemus hermannunezi is only known from the type locality ( Fig. 5 View FIGURE 5 ).
Natural history. We know little about the biology of L. hermannunezi . It is diurnal and inhabits mountainous areas of the Patagonian steppe with short (<0.5 m), bushy vegetation growing on sandy-gravel substrata. This vegetation is used almost exclusively for refuge and on several occasions individuals entering vegetation were found buried in loose sand at the base of the shrub. Nothing is known about its diet or reproductive mode. Individuals were never found under or basking on rocks. The vegetation in the type area is characterized by Acaena pinnatifida, Agrostis serranoi, Arenaria serpens, Berberis empetrifolia, Chaetanthera lycopodioides, Cyperus reflexus, Epilobium nivale, Erigeron andicola, Gaultheria pumila, Geranium sessiliflorum, Haplopappus diplopappus, Heliotropium geissei, Hypochaeris spp., Plagiobothrys myosotoides, Plantago lanceolata, Poa sp., Rumex acetocella , Spergularia rubra , Trifolium spp., and Viola subandina (M. Molina-Montenegro, pers. comm.).


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Liolaemus hermannunezi
| Pincheira-Donoso, Daniel, Scolaro, Alejandro & Schulte, James A. 2007 |
Liolaemus (Eulaemus) rothi Pincheira-Donoso and Núñez (2005:99)
| Pincheira-Donoso and Nunez 2005: 99 |
Liolaemus rothi
| Donoso-Barros 1974: 287 |