
Dematochroma humboldtiana ( Heller, 1916 )
|
publication ID |
https://doi.org/ 10.5281/zenodo.205363 |
|
DOI |
https://doi.org/10.5281/zenodo.6186551 |
|
persistent identifier |
https://treatment.plazi.org/id/03AB87CB-761F-4C40-F2EA-2AF842CFFE71 |
|
treatment provided by |
Plazi |
|
scientific name |
Dematochroma humboldtiana ( Heller, 1916 ) |
| status |
|
Dematochroma humboldtiana ( Heller, 1916)
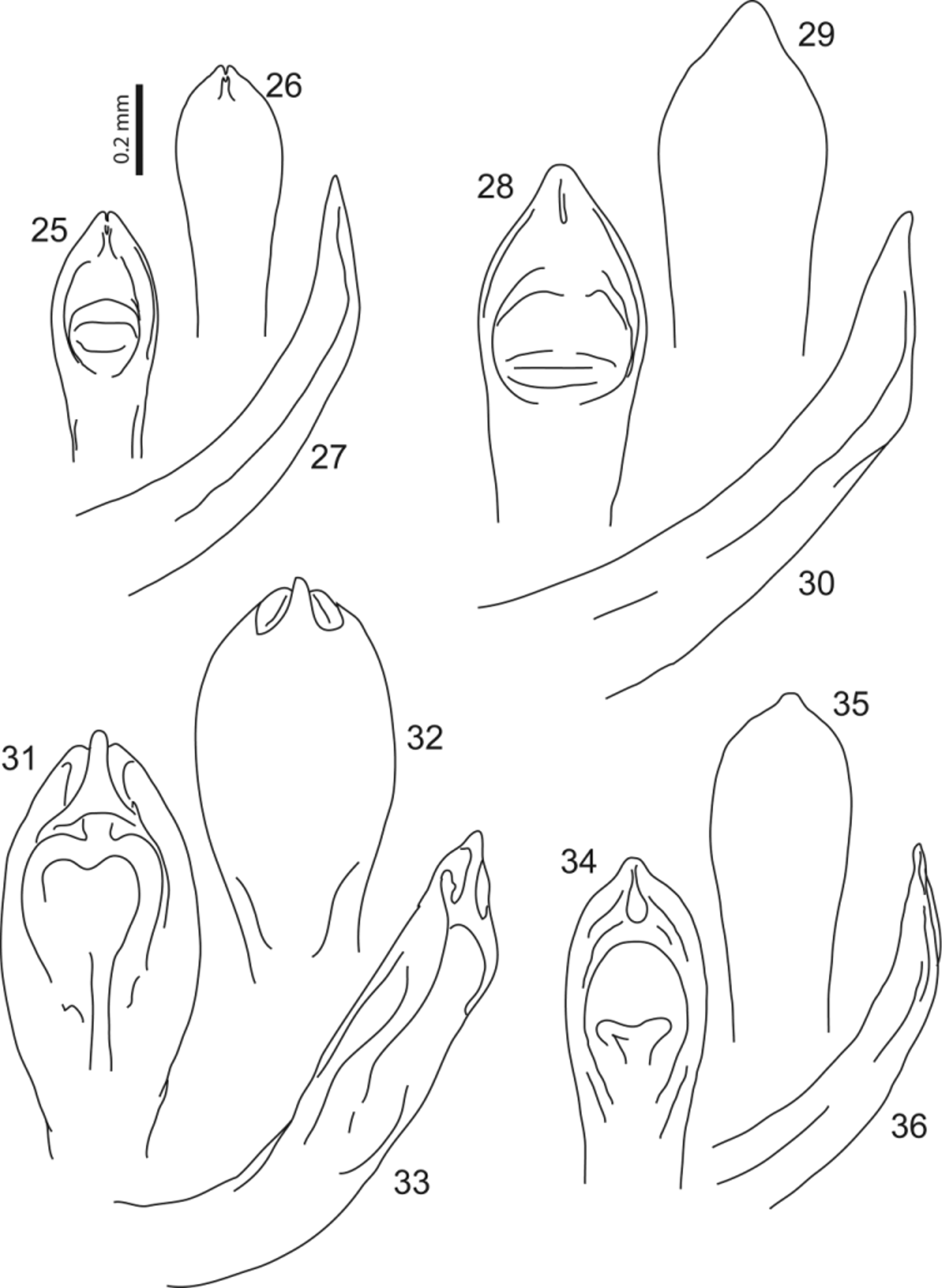
( Figs 2 View FIGURES 1 – 9 , 34–36 View FIGURES 25 – 36 )
Thasycles humboldtiana Heller, 1916:301 .
Dematochroma humboldtiana: Jolivet et al., 2007a:40 .
Type material. Province Sud: Lectotype, 13 ( Fig. 2 View FIGURES 1 – 9 ): Mt. Humboldt, 1100m., 17 September 1911, F. Sarasin & J. Roux leg. [pale blue label], humboldtiana Typus [red label], Staatl. Museum für Tierkunde, Dresden [white label], Dematochroma humboldtianus Heller [white label] (MfT). Paralectotype, 13: Cotypus [red label], Mt. Humboldt, 1100m., 17 September 1911, F. Sarasin & J. Roux leg. [pale blue label] ( NMB).
Other material examined. Province Sud: 13: Cotypus [red label], Mt. Humboldt, Gipfel 1600m., 18 September 1911, F. Sarasin & J. Roux leg. [pale blue label], Thasycles humboldtianus m. Det. K. M. Heller 1915 [white label] (MfT); 13: Mont Kouakoué, 21.95758ºS 166.53830ºE, 1315m, 17 March 2008, J.A. Jurado-Rivera leg. (IBE-JGZ).
Dematochroma humboldtiana (Heller) was described from an unknown number of specimens collected on Mount Humboldt, the second highest mountain in New Caledonia (1618 m), in the mountain range south east of Grande Terre, on 17 September 1911 ( Heller 1916). Of the two syntypes, currently in the MfT and NMB collections, that in the former is designated here as lectotype, and the other as paralectotype. The original description of this species was sufficiently detailed for the recognition of the species and it is complemented here with the description of the median lobe of the aedeagus, while the female still remains unknown.
Male genitalia: Penis ( Figs 34–36 View FIGURES 25 – 36 ) regularly curved in lateral view; weakly, but gradually narrowing towards pointed apex in sagittal plane. Apical third of penis progressively widening slightly before ostium in dorsal view, reaching maximal width (1.5x wider than width at middle) 2/3 before apical end of ostium; ostium egg-shaped, with narrower end basally; sides curved, gradually converging towards apex; apex of penis with blunt round apical short tooth, 0.25x as wide at base as maximal width of penis, 0.5x as long as wide at base. Without prominent ventral features, except for very shallow and wide short median depressed area preapically.
Distribution. This species must be rare or rather localized. Jolivet et al. (2007a) in their second study on New Caledonian Eumolpinae did not find other specimens besides those included in the type series from Mt. Humboldt. The discovery of the species in Mont Kouakoué merely expands its area of distribution to the nearby massif, southeast from Humboldt by less than 15km, across the Ni River valley. Nonetheless, these two massifs probably constitute a single biogeographic unit, since other high altitude leaf beetles such as Cazeresia montana Jolivet et al., 2005 , or Zira nitens Reid and Smith, 2004 , only occur on both mountain peaks (pers. obs.). The narrow geographic endemism of these species, occupying island-like environments within an island is suggestive of strong ecological specialization. This could be perhaps related to host associations with plants presenting this same relictual pattern. However, the purported host plants of both Cazeresia as well as Zira , Dracophyllum involucratum and Myodocarpus spp., respectively ( Jolivet et al. 2005), grow on ultramafic soils in other parts of southern Grande Terre ( Oliver 1952; Anonymous 2011), where these beetle species have not been found to date. However, host specialization coupled with climatic change, e.g. range contractions driven by Quaternary climate change cycles, could be responsible for these species relict distributions. Alternatively, the hypothesized restricted ranges of D. humoldtiana and other leaf beetles in New Caledonia could be later stages of natural biodiversity cycles resulting from evolutionary and ecological interactions among species, as explained by the taxon cycle or taxon pulse hypotheses ( Wilson 1959; Erwin 1981).

| NMB |
Naturhistorishes Museum |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Dematochroma humboldtiana ( Heller, 1916 )
| Gómez-Zurita, Jesús 2011 |
Dematochroma humboldtiana: Jolivet et al., 2007a :40
| Jolivet 2007: 40 |
Thasycles humboldtiana
| Heller 1916: 301 |