
Geocapromys caymanensis, Morgan & Macphee & Woods & Turvey, 2019
|
publication ID |
https://doi.org/ 10.1206/0003-0090.428.1.1 |
|
persistent identifier |
https://treatment.plazi.org/id/03AA87B0-FFFD-FF93-FF6A-16F9FEF3FE05 |
|
treatment provided by |
Carolina |
|
scientific name |
Geocapromys caymanensis |
| status |
sp. nov. |
Geocapromys caymanensis , new species
Figures 19–24 View FIG View FIG View FIG View FIG View FIG View FIG
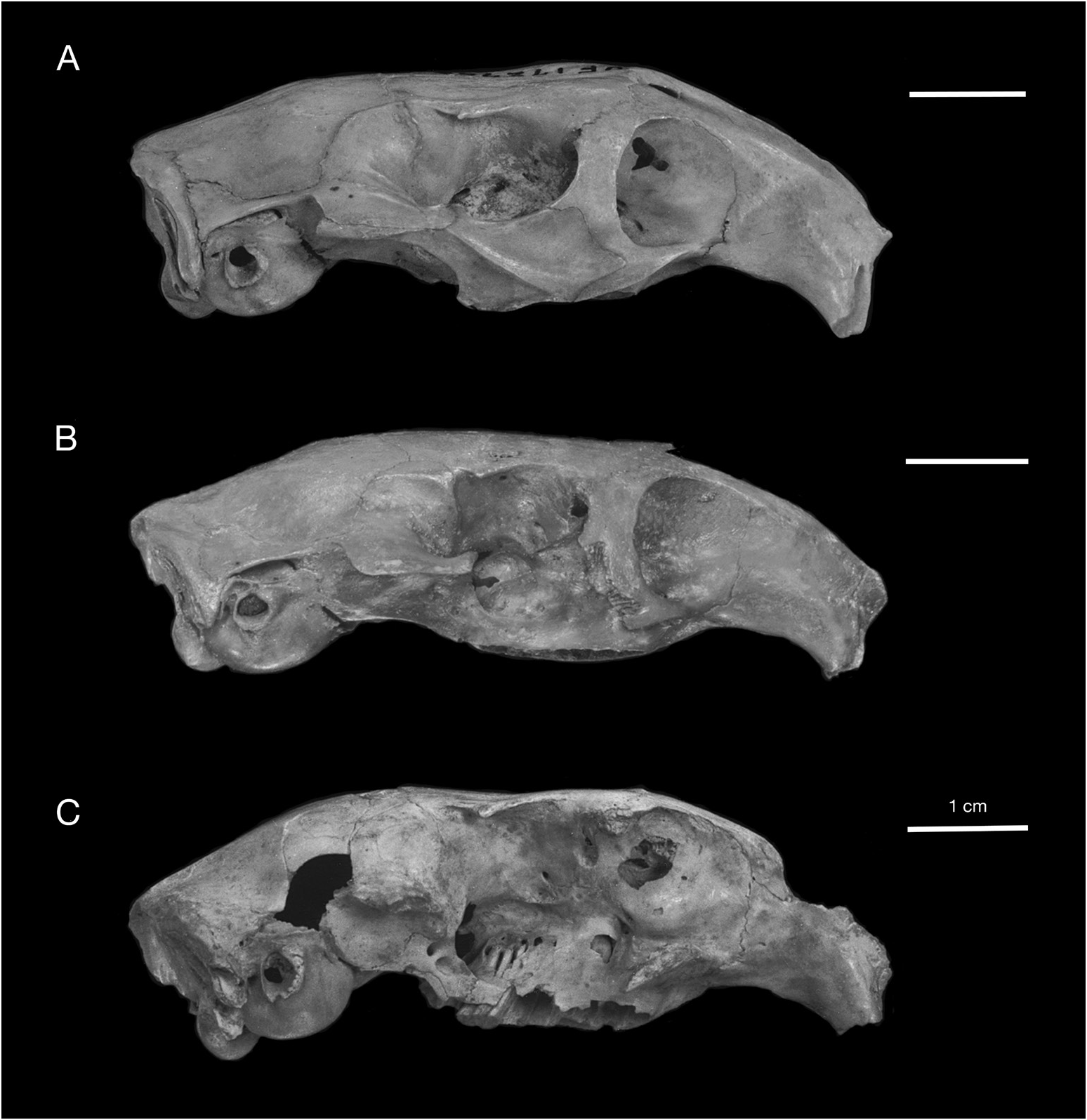
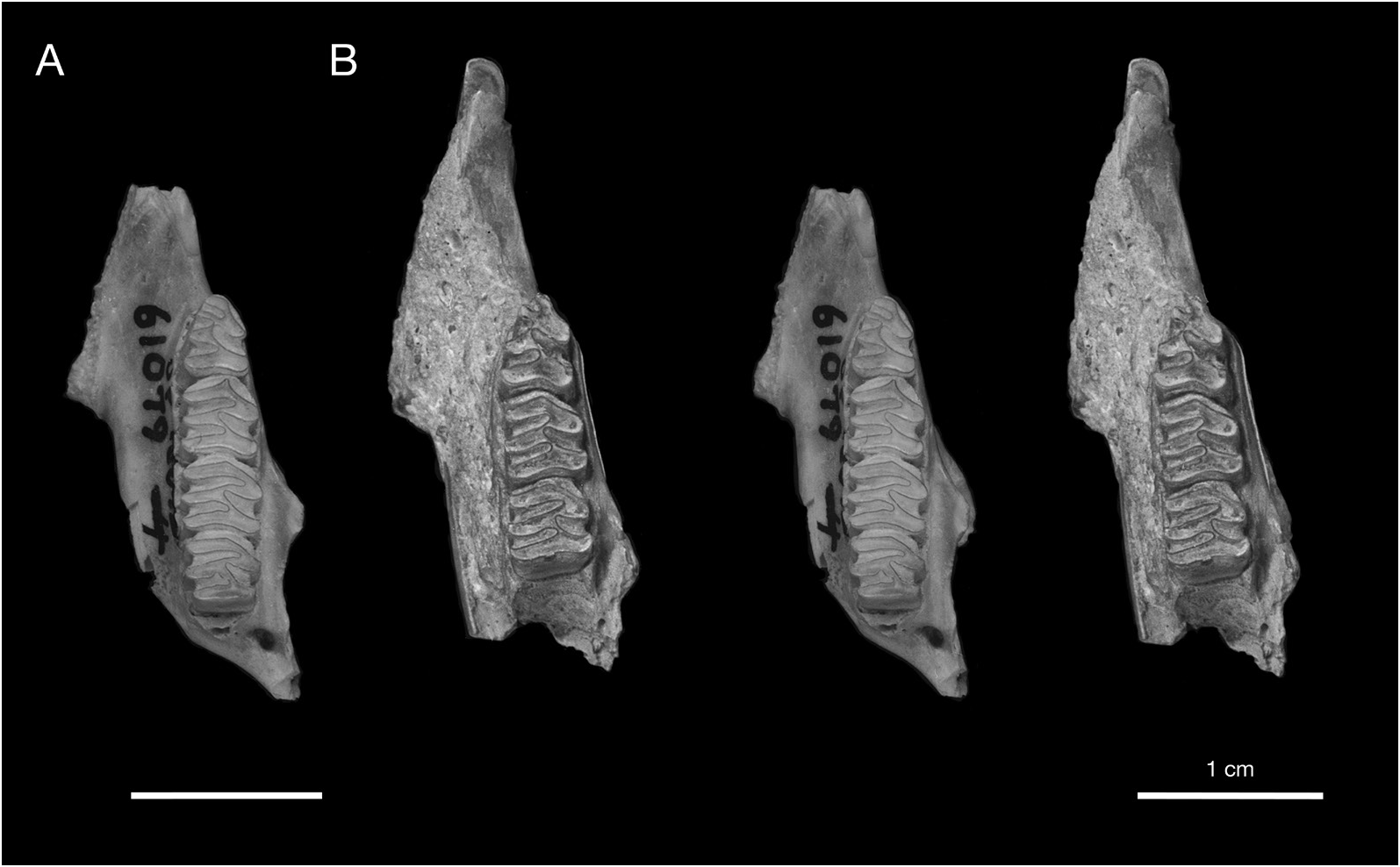
HOLOTYPE: UF 21388 , nearly complete skull with right M3, lacking the left premaxilla, both nasals, and portions of the left and right zygomatic arches (figs. 19C, 20C).
TYPE LOCALITY: Patton’s Fissure, near Spot Bay, northern coast of Cayman Brac, Cayman Islands. The holotype was recovered from layer 7 (120–140 cm below the surface), which has been radiocarbon dated at 13,230 ± 135 yr BP.
ETYMOLOGY: The name caymanensis refers to the Cayman Islands.
AGE: Late Pleistocene–Holocene (see Radiocarbon Dating).
DISTRIBUTION: Known only from Cayman Brac and Grand Cayman.
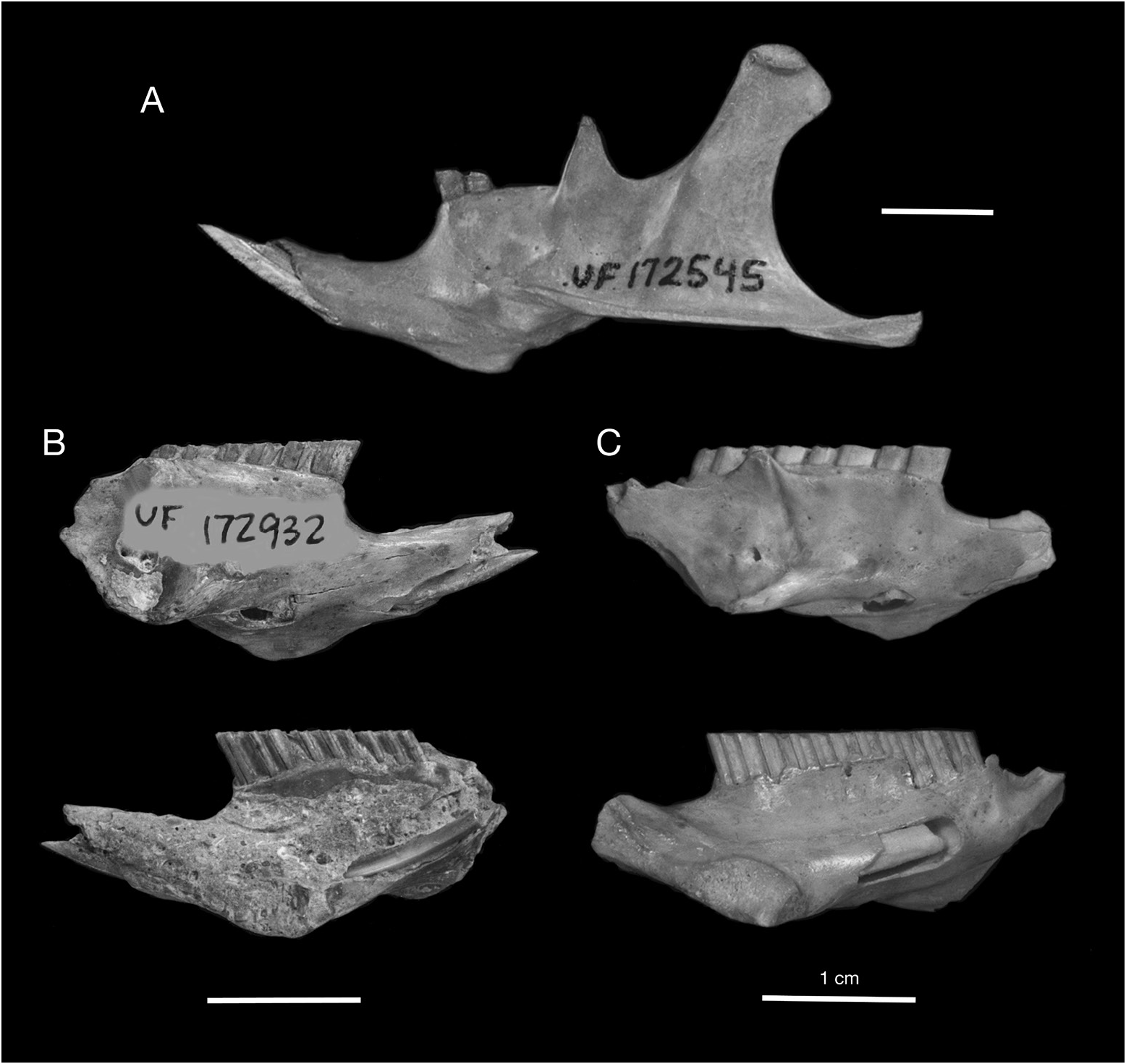
REFERRED SPECIMENS: Cayman Brac: Fig Tree Cave: UF 172761, edentulous mandible. Hutia Cave: UF 172756, nearly complete edentulous skull, lacking nasals, left jugal, and left auditory bulla. Patton’s Fissure: UF 18835, partial skull with left M3, lacking nasals, anterior portion of frontals, left zygomatic arch, and braincase; UF 18836, partial skull, lacking rostrum, teeth, and zygomatic arches; UF 18768, 21358, partial skulls with associated mandibles; UF 21365, partial skull lacking teeth, nasals, zygomatic arches, and braincase; UF 21389, braincase; UF 21398, rostrum with left M2–M3 and right M1–M3; UF 61098, partial skull with left and right M3 and anterior portion of frontals, lacking nasals, zygomatic arches, and braincase; UF 61084, 61086–61089, rostra with edentulous palates; UF 61083, 61090, rostrums; UF 17025, 18553, 18768, 18833, 18836, 21390, 21392, 61099, 61100, partial skulls; UF 61093, palate with right M1–M3; UF 61094, maxilla with M1– M3; UF 61095, palates with left and right M2– M3; UF 21403, associated edentulous maxilla and mandible; UF 61058, 61059, 61078, 61079, mandibles with p4–m3; UF 18821, mandible with p4–m3; UF 61052, mandible with p4–m1; UF 61053, 61060–61062, mandibles with m1– m3; UF 61054, 61063, 61064, mandibles with m1–m2; UF 61049, 61050, 61055, 61065, 61066, mandibles with m2–m3; UF 21403, 61048, 61051, 61057, 61080, 61082, edentulous mandibles; UF 18651, 18652, 18670–18678, mandibles. Peter Cave: UF 22856, partial skull lacking teeth, nasals, zygomatic arches, and braincase; UF 23707, partial skull. Pollard Bay Cave: UF 17025, 18553, partial skulls; UF 18542, mandible; UF 17048, mandible with i1. Spot Bay Cave: UF 61272, partial skull with left M1–M2 and right P4–M3; UF 61273, partial skull with right M1; UF 21674, palate with left M1–M3 and right M1; UF 172780, partial skull with left otic region; UF 172768, edentulous palate; UF 61256, mandible with i1, p4–m3; UF 61257, 172769, mandibles with p4–m2; UF 172775, mandible with m1–m3; UF 172770, mandible with i1, m1–m2; UF 172776, mandible with i1, m2–m3; UF 61260, 61261, 61269–61271, 172772–172774, 172777– 172779, edentulous mandibles. Grand Cayman: Agouti Cave: UF 172883, mandible with i1, m1; UF 172884–172886, edentulous mandibles. Barn Owl Cave: UF 23742, mandible with p4–m2; UF 23741, mandible with m2–m3; UF 23738, 23739, 23745, edentulous mandibles. Bodden Cave: UF 23749, partial maxilla with P4; UF 23747, 23748, edentulous palates. Chisholm Cow Well: UF 172831, 172832, humeri; UF 172833, 172834, ulnae; UF172835, tibia. Crab Cave: UF 23729, partial maxilla with P4; UF 23722, mandible with p4–m1; UF 23712–23721, 23723–23727, edentulous mandibles. Dolphin Cave: UF 172941, palate with left P4–M2 and right M1– M2; UF 172858, palate with left M1; UF 172913, 172930, 172931, edentulous palates; UF 172859, 172914, 172916, 172932, mandibles with p4–m3; UF 172850, 172852, mandibles with p4–m2; UF 172851, 172936, 172937, mandibles with m1– m3; UF 172933, mandible with m1–m2; UF 172917, mandible with m1; UF 172853–172857, 172915, 172918, 172919, 172922, 172923, 172925, 172935, 172938, 172942, 172943, 411278, 411279, edentulous mandibles. Furtherland Farms Cow Well: UF 172544, nearly complete edentulous skull, lacking nasals, left premaxilla, and jugals; UF 172808, partial skull; UF 172805, mandible with i1, p4–m3; UF 172806, mandible with i1, p4–m2; UF 172807, mandible with i1, p4–m1; UF 172545, associated mandibles with left i1, p4 and right i1; UF 172546, edentulous mandible. Miller’s Cave: UF 172903, edentulous mandible; UF 172904, 17290, humeri. Old Man Cave: UF 23743, partial edentulous skull with maxillae and frontals (juv.); UF 23745, 23746, 172903, edentulous mandibles. Queen Elizabeth II Botanic Park: UF 172949, palate with left and right P4. Tadarida Cave: UF 172839, edentulous mandible.
DIAGNOSIS: Geocapromys caymanensis is a small species of Geocapromys , slightly larger than G. ingrahami , similar in size to G. thoracatus , and smaller than G. brownii and G. columbianus . The most distinctive feature of G. caymanensis is the strong anterior convergence of the upper toothrows, with left and right P4 separated by less than 2 mm anteriorly and medially. This anterior convergence gives the upper toothrows of G. caymanensis a gentle but noticeable curvature from anterior to posterior. The dorsal surface of the frontals is comparatively narrow and essentially parallel sided from the nasofrontal suture posteriorly to the frontoparietal suture. The anterior portion of the frontals shows an incipient inflation from the orbit anterior to the nasofrontal suture, particularly on the lateral surface. The postorbital processes are weak and bluntly triangular. The dorsal process of the maxilla is vertical and broad anteroposteriorly. The lateral jugal fossa on the zygomatic arch is comparatively small and narrow. The posteroventral edge of the jugal lacks a spine. The zygomatic arch in not rotated ventrally. In lateral view, the ventral surface of the zygomatic arch is level with the alveolar margin of the toothrow. The auditory bullae are rounded and short anteroposteriorly, but show moderate inflation.
DESCRIPTION: The description of Geocapromys caymanensis is based primarily on one nearly complete skull (the holotype, UF 21388) and three partial skulls (UF 18835, 18836, 21365) from Patton’s Fissure on Cayman Brac, and a nearly complete but edentulous skull (UF 172544) from the Furtherland Farms Cow Well on Grand Cayman (figs. 19, 20). Three of the skulls preserve M3 but are otherwise edentulous. Geocapromys has unrooted, ever-growing, highcrowned (hypseledont) teeth that are not firmly rooted in the skull, which results in most of the teeth falling out of the skulls and jaws upon death. Large samples of isolated teeth are often found in fossil sites that contain Geocapromys . The descriptions and illustrations of the teeth are based on other specimens listed above.
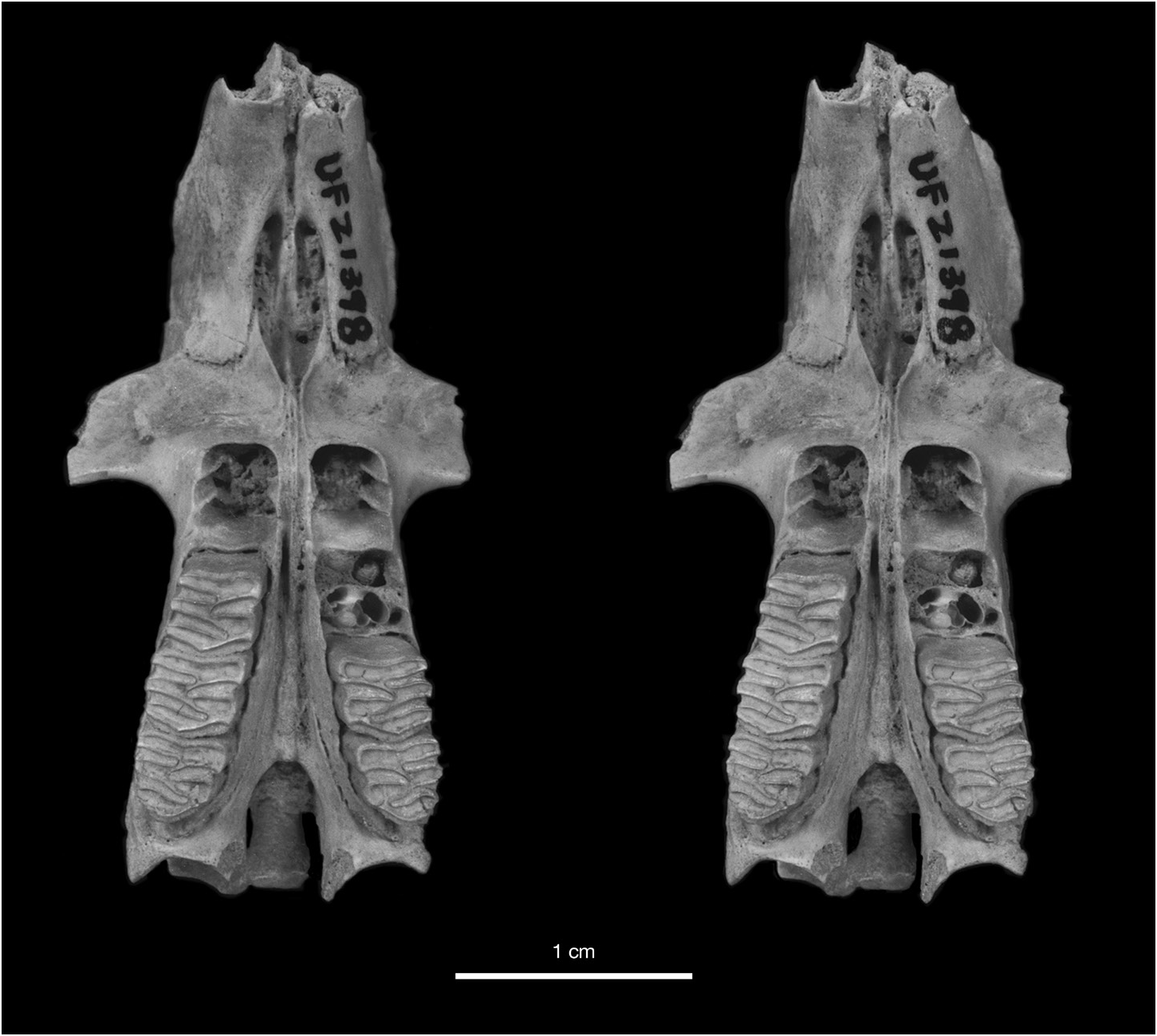
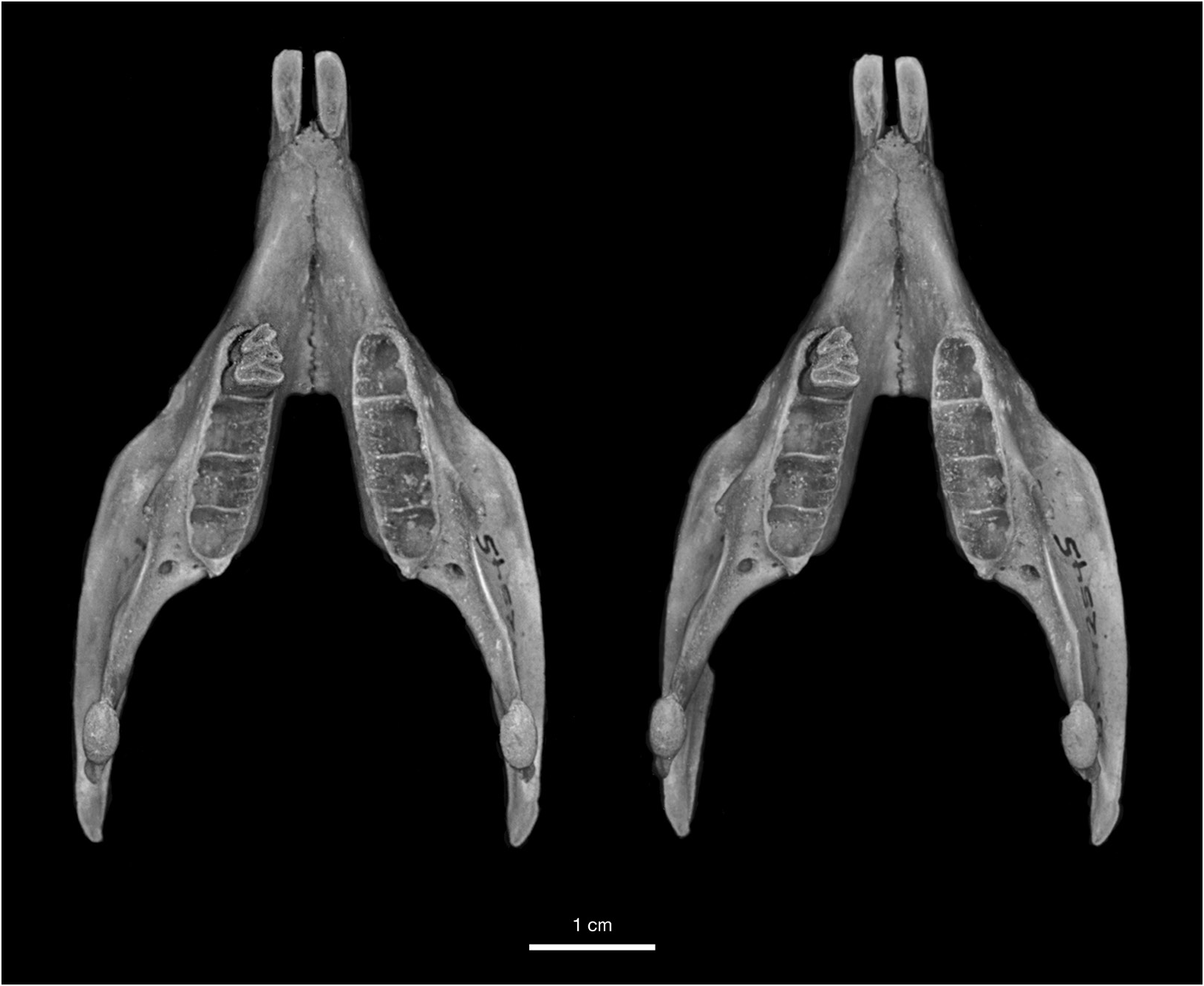
The most distinctive feature of Geocapromys caymanensis is the extreme anterior convergence of the upper toothrows (figs. 19, 21). The left and right P4 nearly meet at the anterior margin of the toothrow. The width of the palate remains very narrow for the entire length of the P4s, with left and right toothrows separated by less than 2 mm at the alveolar margin between P4 and M1. Based on measurements of palatal width (table 6), the palate is actually slightly narrower at the alveolar margin between P4 and M1 (1.7 mm, mean of five individuals) than anterior to P4 (1.9 mm, mean of five individuals). Posterior to the middle of M1, the toothrows begin to diverge laterally, with the distance between toothrows reaching a width of about 5 mm at the posterior margin of the palate. This is similar to the posterior width of the palate in the two other small species of Geocapromys , G. ingrahami and G. thoracatus . Because of their strong anterior convergence at the level of P4 and rather sharp divergence posterior to M1, the upper toothrows have a gentle but noticeable curvature from anterior to posterior, particularly obvious on the lateral alveolar margin.
Another diagnostic character of Geocapromys caymanensis is the narrow, parallel-sided frontals on the dorsal surface of the skull (fig. 19). Compared to other species of Geocapromys , the frontals of G. caymanensis are especially narrow anteriorly, from the level of the least interorbital breadth anteriorly to the nasofrontal suture, and are only slightly broader posteriorly between the postorbital processes and the frontoparietal suture. Measurements that reflect the narrow frontals are the breadth of the frontals both anterior to the supraorbital processes (interorbital breadth) and posterior to the supraorbital processes (postorbital breadth). These frontal measurements are narrower than the same measurements in all other species of Geocapromys , even though G. caymanensis is not the smallest species in the genus (tables 6, 7). Although narrow transversely, the anterior frontals in the Cayman Geocapromys are moderately inflated, from dorsal to the anterior edge of the orbit anteriorly to the nasofrontal suture. This anterior frontal inflation is primarily obvious laterally along the internal margin of the orbit and dorsal to the superior process of the maxilla, but not along the midline where the frontals are essentially flat. The widest portion of the frontals occurs at the level of the postorbital processes, which are small and bluntly triangular. Compared to other Geocapromys , the postorbital processes are weak in G. caymanensis . The temporal crests are low, rather weak, U-shaped, and do not meet to form a sagittal crest; they are separated by 3–4 mm on the posterior surface of the parietals, just anterior to their connection with the nuchal crest.
The dorsal process of the maxilla is preserved in only one skull of G. caymanensis (holotype; fig. 19C), in which this process is vertical and relatively broad anteroposteriorly. The lateral jugal fossa (a character present on the posteroventral margin of the zygomatic arch ventral to the orbit in capromyines) is preserved in only one of the Geocapromys skulls from Cayman Brac (UF 18835), in which this fossa is narrow and its ventral margin lacks a jugal spine. In lateral view, the zygomatic arch in this skull of G. caymanensis is not noticeably downturned or rotated ventrally. The ventral edge of the zygomatic arch is only slightly inclined posteroventrally and is on essentially the same level as the alveolar margin of the toothrow. In ventral aspect, the pterygoid region is constricted anteroposteriorly. This compressed pterygoid region is indicative of a rather foreshortened braincase that is also noticeably downturned ventrally. The auditory bullae are comparatively short, rounded, and somewhat inflated. In posterior view, the auditory bullae extend ventrally to the occipital condyles.
MORPHOMETRICS: As for Capromys , the largest available measurement series for the Cayman Geocapromys sample is the mandibular alveolar toothrow length (Grand Cayman, n = 41; Cayman Brac, n = 30). Although Geocapromys skulls from Grand Cayman and Cayman Brac are otherwise morphologically very similar, statistically detectable size differences are present between samples from these two islands (Welch two-sample t-test not assuming equal variance: Grand Cayman, mean = 15.25 mm; Cayman Brac, mean = 16.52 mm; p <0.001). Sample series from each island were therefore compared separately with morphometric data for other extant and extinct Geocapromys populations measured from museum collections: G. brownii (n = 17, mean = 19.11 mm), G. columbianus (n = 40, mean = 18.76 mm), G. ingrahami (sample comprising both recent specimens from East Plana Cay, and Quaternary fossil specimens from Abaco, Crooked Island, and Exuma: n = 57, mean = 16.12 mm), and G. thoracatus (n = 12, mean = 14.72 mm) (appendix 2). Our Grand Cayman Geocapromys sample has a significantly smaller mandibular alveolar toothrow length compared to G. brownii (p <0.001), G. columbianus (p <0.001) and G. ingrahami (p <0.001), and a significantly greater mandibular
alveolar toothrow length compared to G. thoracatus (p = 0.016). Our Cayman Brac Geocapromys sample has a significantly smaller mandibular alveolar toothrow length compared to G. brownii (p <0.001) and G. columbianus (p <0.001), a significantly greater mandibular alveolar toothrow length compared to G. thoracatus (p <0.001), and does not differ statistically in size from G. ingrahami (p = 0.089).
MORPHOLOGICAL COMPARISON WITH OTHER SPECIES OF GEOCAPROMYS : Geocapromys is the
most widespread genus in the Capromyinae , known as either a living animal or from historical museum specimens or fossils from the Cayman Islands, Bahamas, Cuba, Jamaica, and Little Swan Island. Geocapromys caymanensis is a rather small species of Geocapromys , comparable in size to the smallest of the three living species, G. ingrahami from the Bahamas (tables 6, 7). Among the three extant species of Geocapromys , G. caymanensis is most similar to the Bahamian hutia G. ingrahami and least similar to the Jamaican hutia G. brownii .
Geocapromys ingrahami View in CoL survives today as a wild population on a single island, East Plana Cay in the southern Bahamas, with translocated populations also established on Little Wax Cay and Warderick Wells Cay (Clough, 1972; Turvey et al., 2017). Two extinct subspecies have been described based on fossils from the Bahamas: G. ingrahami irrectus from Crooked Island, with referred samples from Eleuthera and Long Island, and G. ingrahami abaconis from Abaco (Lawrence, 1934). Both subspecies were distinguished from the living G. ingrahami View in CoL primarily on the basis of their larger size (e.g., longer alveolar toothrows). Numerous additional extinct populations of G. ingrahami View in CoL have been reported since, mostly from islands on the Great Bahama Bank, although these samples were not referred to either of the two extinct subspecies ( Morgan, 1989a). Geocapromys caymanensis overlaps with G. ingrahami View in CoL in many of the most important cranial measurements indicative of size, including total length and condylobasal length of the skull, and breadth of the skull at the auditory meatus. The general size similarity in Geocapromys View in CoL individuals from these two geographically separate island groups may be explained by the relatively small land area of islands in both regions compared to the much larger islands of Cuba and Jamaica; this may have resulted in common constraints on maximum body size attainable in both regions (cf. Burness et al., 2001). The Bahamian and Cayman species of Geocapromys View in CoL differ in the degree of anterior convergence of upper toothrows, which are highly convergent in G. caymanensis , and convergent but less extreme in G. ingrahami View in CoL . Measurements of the anterior palatal width show that the toothrows are more strongly convergent anteriorly in these two species than in G. brownii View in CoL and G. thoracatus View in CoL (table 6). The frontals of G. ingrahami View in CoL are slightly broader than in G. caymanensis , which has narrower frontals compared with all other species in the genus. G. ingrahami View in CoL shows no evidence of the anterior frontal inflation that is characteristic of G. caymanensis , and also has more prominent, sharply triangular postorbital processes compared to the small, blunt processes of G. caymanensis . The dorsal process of the maxilla is vertical in both G. caymanensis and G. ingrahami View in CoL but is noticeably broader in the Cayman species. These two species share several characters of the zygomatic arch that may also indicate a close phylogenetic relationship, including the narrow jugal fossa, lack of a jugal spine, and the minimal ventral rotation of the entire zygomatic arch with the ventral edge of the arch level with the alveolar margin of the toothrow. Both G. ingrahami View in CoL and G. caymanensis also have a foreshortened braincase that is downturned ventrally. The short braincase is reflected ventrally in the constricted pterygoid regions of these two species. G. ingrahami View in CoL has comparatively larger auditory bullae that are more inflated than in the Cayman species.
Geocapromys caymanensis differs in size and most diagnostic cranial characters from the largest living species in the genus, G. brownii View in CoL from Jamaica. Measurements in tables 6 and 7 reveal no overlap in cranial measurements between the smaller G. caymanensis and the larger G. brownii View in CoL . The strong anterior convergence of the upper toothrows in G. caymanensis is not observed in G. brownii View in CoL . The characters of the dorsal surface of the frontals also differ considerably between these two species. In G. brownii View in CoL , the frontals are very broad and strongly inflated anterior to the postorbital processes, especially laterally, whereas the frontals are quite narrow and constricted posterior to the postorbital processes. Overall, the frontals in G. brownii View in CoL are much broader anterior to the postorbital processes than posterior to these processes (table 6). In G. caymanensis , the frontals are comparatively narrow and almost parallel sided, exhibit only a minor degree of anterior inflation, and are slightly broader posterior to the postorbital processes. G. brownii View in CoL has rather weak V-shaped temporal crests that meet to form a prominent sagittal crest on the parietals about midway between the frontal and occipital sutures. The temporal crests are also weak in G. caymanensis , but tend to be more U-shaped, converge (but do not meet) much farther posteriorly on the parietals, and do not form a sagittal crest. G. caymanensis has a foreshortened and ventrally downturned braincase compared to G. brownii View in CoL , in which the braincase is more elongated. The comparative length of the braincase is especially obvious in the pterygoid region, in which the distance between the posterior edge of the toothrow and the anterior edge of the auditory bulla is much shorter in G. caymanensis than in G. brownii View in CoL . The dorsal process of the maxilla is vertical in G. caymanensis , whereas this process is more posteriorly oriented in G. brownii View in CoL . The posterior orientation of the dorsal process of the maxilla in G. brownii View in CoL is related to the overall ventral rotation of the zygomatic arch, with the ventral edge of the arch located markedly ventral to the alveolar margin and occlusal surface of the upper teeth. The zygomatic arch is not rotated ventrally in G. cay- manensis; the ventral edge of the arch is instead located approximately level with the alveolar margin of the toothrow. G. brownii View in CoL has a large, broad jugal fossa with a prominent jugal spine located on the posteroventral margin of the jugal; a jugal spine is absent in G. caymanensis , and the jugal fossa is narrow. The auditory bullae are larger and more elongated in G. brownii View in CoL , versus shorter and rounded in G. caymanensis .
The Little Swan Island hutia, G. thoracatus , is somewhat larger than G. caymanensis in most cranial measurements (table 6), and shares many morphological features with the larger Jamaican G. brownii , to which G. thoracatus appears to be closely related ( Morgan, 1985). This species is now extinct but survived on Little Swan Island until the 1950s, and is represented by skins, skulls, and skeletons in several museum collections ( Morgan, 1989b; Tonge, 2014). Unlike G. caymanensis , G. thoracatus does not exhibit a strong anterior convergence of the upper toothrows. This species has a small spinous process that projects 1–2 mm beyond the posterior palatal margin along the midline; this process is not observed in any other species of the genus, including G. caymanensis . One of the most char- acteristic features of G. thoracatus compared to its congeners is its comparatively small teeth. The frontals are broad in G. thoracatus , especially posterior to the postorbital processes, and there is no evidence of anterior inflation of the frontals, compared with G. caymanensis , which has narrow, parallel-sided frontals with a minor degree of inflation anteriorly. Features of the dorsal process of the maxilla and zygomatic arch of G. thoracatus are similar to those of G. brownii , although less pronounced. G. thoracatus has a broad, posteriorly oriented dorsal process of the maxilla, a broad jugal fossa with a well-developed jugal spine, and a ventrally rotated zygomatic arch, all of which are quite distinct from G. caymanensis ; the Cayman species instead has a vertical dorsal process of the maxilla, a narrow jugal fossa lacking a jugal spine, and an absence of ventral rotation of the zygomatic arch. G. thoracatus has longer and narrower auditory bullae than G. caymanensis , which has shorter, rounded, and more inflated bullae.
Four extinct species of Geocapromys have been named from Cuban fossil deposits, but only G. columbianus is now considered valid ( Silva Taboada et al., 2007). G. columbianus is not well described or illustrated, with little cranial material having been figured in the literature ( Silva Taboada et al., 2007; Díaz-Franco and Jiménez Vázquez, 2008). In his original description of G. columbianus, Chapman (1892) characterized this species as having strongly convergent upper toothrows, with a measurement of “0.04 in” (= 1.0 mm) between the anterior margins of the P4s. He also noted that the left and right than the Cayman and Cuban species of Geocapalveoli essentially meet between the upper pre- romys. Although not specifically mentioned in molars. Allen (1917) obtained additional fossil the description, the illustration of the holotype specimens of G. columbianus from Cuba and of G. columbianus shows a rather strong curvaconfirmed several of the morphological features ture of the upper toothrows, especially along the of this species reported by Chapman (1892), in lateral margin (Chapman, 1892: fig. 3), a feature particular the strong anterior convergence of also characteristic of G. caymanensis but not the upper toothrows. The Cayman species also quite as pronounced. Chapman (1892) did not has strongly convergent upper toothrows, with provide a measurement of the alveolar length of the anterior margins of the P4s separated by 1.9 the toothrow in the holotype of G. columbianus mm (mean of five individuals; table 6), but this because the palate is broken off posterior to M2. convergence is apparently not quite as extreme as Silva Taboada et al. (2007) presented selected in the Cuban species. The toothrows in G. cay- measurements of Geocapromys columbianus , manensis are also highly convergent between P4 including several cranial measurements of a sin- and M1, separated by only 1.7 mm (mean of five gle skull. The condylobasal length of this skull individuals; table 6); however, the left and right (65.9 mm) is about 12% larger than the only toothrows do not nearly meet between the P4s as skull of G. caymanensis on which this measurein G. columbianus (Chapman, 1892: 314–315, fig. ment could be taken, the holotype (58.4 mm). 3). The strong anterior convergence of the upper The breadth at the auditory meatus in the single toothrows may indicate a close phylogenetic skull of G. columbianus is 28.1 mm, versus a relationship between G. caymanensis and G. mean of 26.4 mm (range of 25.2–27.1 mm) for columbianus . The Bahamian hutia G. ingrahami three specimens of G. caymanensis , about 10% also demonstrates an anterior convergence of smaller than the Cuban species. Silva Taboada et the upper toothrows, although to a lesser degree al. (2007) listed a mean of 16.5 mm (range of 14.4–18.2 mm; 48 specimens) for the alveolar length of the upper toothrow of G. columbianus . Although the mean for the alveolar length of the upper toothrow in five specimens of G. caymanensis is slightly larger at 16.9 mm, the upper limit for the observed range is 17.6 mm, somewhat less than in G. columbianus . Based on the wide range of values, we suspect Silva Taboada et al. (2007) included some juveniles in their sample, whereas we excluded juveniles, thus increasing the mean length of the alveolar toothrow compared with the Cuban sample. Overall, the available measurements suggest that G. columbianus is about 10% larger than the Cayman species. Other cranial characters of G. columbianus that differ from G. caymanensis are: the posterior orientation of the dorsal process of the maxilla, a slight ventral rotation of the zygomatic arch, the presence of a small jugal spine, and the near convergence of the temporal crests on the parietals to form a short low sagittal crest. In contrast, G. caymanensis has a vertical dorsal process of the maxilla, no ventral rotation of the zygomatic arch, no jugal spine, and the temporal crests do not meet.
REMARKS: The species of Geocapromys form a monophyletic group that is distinguished from Capromys by a number of external and cranial characters ( Morgan, 1985; 1989b). External characters that distinguish Geocapromys from Capromys include: shorter tail, reduced first digit on the front foot, and shorter finer fur. Dental and cranial features that characterize Geocapromys are: more procumbent incisors, origin of the upper incisor root capsule high on maxilla above P4, broad vertically or posteriorly oriented dorsal process of the maxilla, 30° inclination of all cheekteeth, and presence of an anteroflexid on p4. Among the three living or recently extinct species of Geocapromys , G. ingrahami is distinguished from G. brownii and G. thoracatus by the following characters that are considered derived for G. ingrahami : more convergent upper toothrows, constricted pterygoid region, shortened and more inflated braincase, and inflated auditory bullae. The extinct taxa of Geocapromys from Cuba and the Cayman Islands possess most of the derived characters present in G. ingrahami , although the expression of these characters varies significantly between species. The shared possession of these derived characters suggests that G. caymanensis and G. columbianus are probably more closely related to G. ingrahami than they are to G. brownii or G. thoracatus , and these species were previously associated as the ingrahami species group within Geocapromys ( Morgan, 1985) .
From a biogeographic standpoint, the extinct G. columbianus from Cuba represents a plausible source population for G. caymanensis . Only G. columbianus and G. caymanensis exhibit extreme anterior convergence of the upper toothrows, which is almost certainly a derived feature indicating a close relationship between these species. Considering Cuba’s central location between the Bahamas to the north and the Cayman Islands to the south, as well as the long history and evolutionary diversity of capromyine rodents in Cuba, it seems most likely that G. columbianus or a precursor probably gave rise to both G. caymanensis and G. ingrahami sometime prior to the Late Pleistocene.
Geocapromys View in CoL occurs in almost every cave deposit excavated in both Cayman Brac and Grand Cayman (see list of Referred Specimens). No fossils of Geocapromys View in CoL are known from Little Cayman, although only limited fossil exploration has been conducted on that island. As in his initial assessment of the status of the then-undescribed Nesophontes View in CoL populations from Grand Cayman and Cayman Brac, Morgan (1994a) proposed that Geocapromys View in CoL populations from these two islands varied in size, and should therefore be interpreted as representing two distinct species. We support Morgan’s (1994a) observation by revealing statistically detectable size differences between Geocapromys View in CoL samples from Grand Cayman and Cayman Brac, but, as with the allopatric Cayman Nesophontes View in CoL populations, these samples are otherwise morphologically indistinguishable and lack any island-specific unique morphological synapomorphies. We are therefore reluctant to taxonomically differentiate the two allopatric Cayman Geocapromys View in CoL populations in the absence of further information about their phylogenetic relationships, and we encourage future investigation of the evolutionary history of these populations using ancient DNA techniques.

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Geocapromys caymanensis
| Morgan, Gary S., Macphee, Ross D. E., Woods, Roseina & Turvey, Samuel T. 2019 |
Geocapromys
| Chapman 1901 |
Geocapromys
| Chapman 1901 |
Geocapromys
| Chapman 1901 |
Geocapromys
| Chapman 1901 |