
Tropidurus sertanejo, Carvalho & Sena & Peloso & Machado & Montesinos & Silva & Campbell & Rodrigues, 2016
|
publication ID |
https://doi.org/ 10.1206/3852.1 |
|
persistent identifier |
https://treatment.plazi.org/id/03AA7F67-5B2F-2117-FE53-13A42CFE0A3A |
|
treatment provided by |
Carolina |
|
scientific name |
Tropidurus sertanejo |
| status |
sp. nov. |
Tropidurus sertanejo , n. sp.
Figures 1 View FIGURE 1 , 2 View FIGURE 2 , 5 View FIGURE 5 –8
HOLOTYPE: MZUSP 104273, adult male from Reserva Particular do Patrimônio Natural Fazenda Pé da Serra , Serra do Arame , Municipality of Ibotirama, State of Bahia, Brazil, (12° 08′ 45.21 S, 43° 03′ 20.83 W, WGS84 system; ~ 507 m above sea level), collected by A.L.G. C., P.L.V. P., and R.M. in 28 July 2013. GoogleMaps
ALLOTYPE: MZUSP 104274, adult female, collected with the holotype (12° 08′ 41.99 S, 43° 03′ 08.32 W, WGS84 system, ~ 516 m above sea level).
PARATYPES: MZUSP 104272, juvenile, collected with the holotype (12° 08′ 41.99 S, 43° 03′ 08.32 W, WGS84 system; ~ 516 m above sea level) by A.L.G.C., P.L.V.P., and R.M. on 28 July 2013. MZUSP 105262 (= RU 6311), adult male, MZUSP 105263 (= RU 6312), adult female, MZUSP 105261 (= RU 6310) and MZUSP 105264–65 (= RU 6313–14), three juveniles, collected in the type locality (12° 08′ 40.06 S, 43° 03′ 23.40 W, WGS84 system; ~ 490 m above sea level) by
A.L.G.C., H.R.S., and R.M. on 22 July 2009. MZUSP 105266–69 (= RU 6353–6356): four juveniles, collected in the type locality (12° 08′ 40.06 S, 43° 03′ 23.40″ W, WGS84 system; ~ 490 m above sea level) by A.L.G.C., H.R.S., and R.M. on 22 July 2009. MZUSP 76048–49, 76055, three adult males, MZUSP 76050-52, three adult females, MZUSP 76046–47, 76053–54, 76056–58, seven juveniles collected in the municipality of Caetité , State of Bahia, Brazil (14° 04′ 17.82 S, 42° 29′ 48.33″ W, WGS84 system; ~ 940 m above sea level), by M.T.R. on 19 September 1991 GoogleMaps .
MORPHOLOGICAL DIAGNOSIS: Tropidurus sertanejo , n. sp., is diagnosed based on a combination of macrostructural characters 7 suggested by Frost et al. (2001) as exclusive to Tropidurus : skull not highly elevated at the level of the orbits; premaxilla not broad; nutritive foramina of maxilla strikingly enlarged; lingual process of dentary present, extending over lingual dentary process of coronoid; angular strongly reduced; medial centrale absent; “flash” marks on undersides of thighs present; circumorbitals distinct from other small supraorbital scales; lateral fringe not developed on both sides of fourth toes; enlarged middorsal scale row absent; and tail terete. Frost et al. (2001) also listed the hemipenis attenuate without apical disks as characteristic of Tropidurus , however the hemipenial morphology of T. sertanejo , n. sp., was not examined.
Tropidurus sertanejo , n. sp., is diagnosed as a member of the T. torquatus group by lacking the enlarged middorsal scale row (well marked in species of the T. spinulosus group, especially in males), by having black thigh flash marks (males of T. spinulosus group have yellow, pale, or white flash marks), and by not being extremely flattened dorsoventrally (as observed in species of the T. semitaeniatus group and, more moderately, in T. bogerti ).
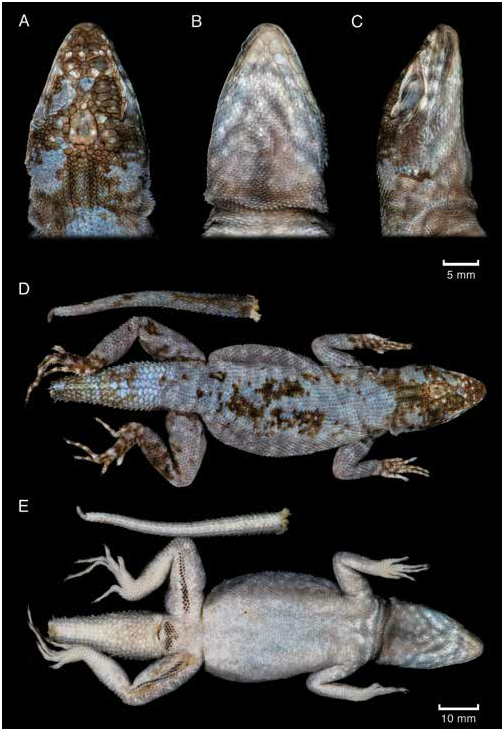
Tropidurus sertanejo , n. sp., lacks granular mite pockets on the lateral neck. The oblique neck fold of the species is covered with imbricate, smooth, mucronate scales, similar to but smaller than temporals and dorsals (fig. 8). The new species has deep, oblique axillary and wide inguinal mite pockets, both coated with unpigmented diminutive granular scales. The bronze head and light brown dorsal body decorated with small pale salmon spots constitute a unique coloration pattern (figs. 2, 4–5). This exclusive combination of macroscopic attributes makes T. sertanejo , n. sp., a rare example of easily diagnosable species within the T. torquatus group.
COMPARISON WITH OTHER SPECIES: Tropidurus sertanejo , n. sp., is the only species of the T. torquatus group lacking mite pockets on the lateral neck and could not be classified according to the eight mite pockets patterns described by Rodrigues (1987). We amended that classification scheme by adding two patterns (I and J) to accommodate T. sertanejo , n. sp., and T. psammonastes (fig. 8; table 2). Tropidurus sertanejo , n. sp., is known to occur in sympatry with only two other forms of the T. torquatus group: T. hispidus and T. aff. etheridgei . However, those species differ considerably with respect to number and morphology of their mite pockets. Tropidurus hispidus has one mite pocket on the lateral neck, one deep and oblique granular axillary mite pocket, and lacks an inguinal pocket. Tropidurus aff. etheridgei has two mite pockets on the lateral neck, and lacks both axillary or inguinal mite pockets, while T. sertanejo , n. sp., lacks a mite pocket on the lateral neck and has both axillary and inguinal pockets well developed and coated with granular scales.
7 Osteological characters were analyzed through digital inspection of the skeleton of the holotype via computed tomography.
The state of conservation of old specimens sometimes precludes accurate identification of the type and number of mite pockets on the lateral neck of specimens. Therefore, checking for the presence of axillary and inguinal granular pockets is an easy way to narrow down the number of species for comparison. Tropidurus sertanejo , n. sp., shares both axillary and inguinal mite pockets exclusively with T. erythrocephalus , T. montanus , and T. mucujensis . These three forms are allopatric with respect to T. sertanejo , n. sp., and have distributions nearly restricted to rocky fields known as campos rupestres, spread over the Espinhaço mountain range, in the states of Minas Gerais and Bahia, Brazil ( Rodrigues, 1987, 1988; Carvalho, 2013). In terms of coloration, T. sertanejo , n. sp., has a bronze dorsal head, distinct from the intense brick-reddish head coloration of T. erythrocephalus . The ventral side of its head is pale salmon, and grades into a dark bronze throat, differing from the orange pigmentation covering the throat and chest of T. erythrocephalus . Tropidurus sertanejo , n. sp., has a dotted dorsal pattern somewhat similar to T. mucujensis , but the former is decorated with pale salmon spots on the dorsum, while the dorsal color pattern in the latter is scattered with sky-blue spots against the dark background of its dorsum and tail. Tropidurus sertanejo , n. sp., also lacks aculeate spines on the lateral neck, a morphological attribute exclusive to T. mucujensis .
DESCRIPTION OF HOLOTYPE: Small species of Tropidurus, SVL 79.92 mm; head subtriangular, length 29% of SVL and width 66% of head length; skull not compressed, not strongly elevated at level of orbits; rostrum not noticeably shortened relative to most other species of Tropidurus ; scales of frontonasal region not imbricating posteriorly, several lenticulate scale organs present (scale organs randomly distributed on other areas of the head); rostral tall, about three times as high as first supralabial, slightly tumescent, contacting first supralabials, first lorilabials, and three postrostrals; 1/2 postrostrals (i.e., right postrostral entire, left divided); nasal single, higher than adjacent scales, separated from rostral by postrostral-lorilabial contact; 5/6 enlarged supralabials followed by 3/3 smaller scales reaching the rictus oris, never contacting subocular; nostril elliptical, occupying about one third of nasal, positioned posteriorly, directed dorsolaterally; 2/2 canthals between nasal and first superciliary; anteriormost canthal separated from supralabials by 1/1 rows of lorilabials and 1/1 rows of loreals; 8/9 laminate superciliary scales weakly produced vertically; 1/1 dorsally keeled preoculars contacting second canthal and 6/5 loreals; 1/1 dorsally keeled elongate suboculars separated from supralabials by one row of lorilabials; palpebrals granular; second row of palpebrals larger, with developed scale organs; 3 rows of supraoculars, oblique internal row with 8/9, medial row with 6/4, external row with 8/8 small scales, the enlarged ones occupying up to two thirds, and two posteriormost internal scales occupying the whole width of the supraocular area; 1/1 rows of small, angulate circumorbitals; 1/1 rows of short semilaminate scales with lenticulate scale organs linearly distributed along their dorsal face separating circumorbitals and superciliaries; interparietal enlarged, about 1.2 times longer than wide; parietal eye visible, positioned medially on the posterior limit of the first third of the interparietal scale; temporals imbricate, keeled, larger than lateral neck scales and smaller than dorsals and parietals, scale organ positioned on the posterior end of the keel or next to the base of a slight mucron, keels more pronounced on upper than lower temporals; ear shaped as inverted keyhole, canal deep, largest infratarsal scales carinate, tricarinate toward fingers and toes; fingers and toes thin, cylindrical, slightly compressed laterally; supradigital lamellae smooth,
rhomboidal, scale organ positioned on the distal end of the scales; infradigital lamellae tricarinate and mucronate, 14/14 under fourth finger, 20/20
under fourth toe, medial careen larger and more projected than laterals; claws long, curved; preaxial scales of forearm strongly keeled and mucronate grading to smooth scales with no or short mucrons on ventral and postaxial surfaces; scales on hind limb decreasing in size toward ventral surface,
24/24 tibial scales, heavily keeled and mucronate;
dorsal body scales large, keeled, mucronate, grading to scales up to 80% smaller at the level of ear opening and neck; rictal, nuchal, postauricular, supraau-
ricular, dorsolateral, longitudinal neck, and antegular fold absent; shallow postauricular depres-
sion present; oblique neck fold well marked and covered with smooth, slightly mucronate, imbricate scales, similar in shape, but smaller than dorsals and temporals, not forming a mite pocket (i.e., lateral neck fold not forming a pocket coated with granular scales; see fig. 8); antehumeral fold present FIGURE 8. Lateral view of the neck of (A) and well marked, coated with imbricate scales simi- Tropidurus psammonastes (AMNH 138852) lar to those on lateral neck; gular fold incomplete and (B) T. sertanejo , n. sp. (holotype MZUSP 104273), illustrating the lateral mite pockets medially; axillary pocket deep and oblique, coated associated with types I and J, respectively. See with diminutive granular scales; inguinal pocket table 2 for details on mite pocket patterns in wide, with granular scales similar to those in the Tropidurus .
axillary pocket; tail compressed, tapering from the end of the first third to become pointed; second half of tail broken, separated from the first half, tip regrown; caudal verticils absent; scales of tail imbricate, keeled, mucronate, up to three times larger than dorsals; middorsal row of caudal scales expanded, laterally deflected, with strong and highly projected keels, forming a caudal crest after the first third of the tail.
COLORATION IN LIFE: Bronze coloration on the dorsal head encompassing the frontonasal, supraocular, and parietal regions. Loreals and lorilabials transition from bronze to irregularly brown-pigmented scales, with pale salmon or cream background toward labials. Labials cream; infralabials slightly lighter than supralabials. Coloration of labials extends posteriorly forming a light facial stripe that crosses the lower temporal region and reaches the preauricular fringe. Inferior portion of preocular and subocular similar in coloration to lorilabials; uppermost, keeled portion of both scales darker. Spot formed by 3–4 pale salmon angulate scales located above the posterior limit of preocular, internally to preocular corner. First and second rows of pale salmon palpebrals form a light ring around the eye, contrasting with the surrounding dark brown granular scales. Upper temporal region light brown, grading into bronze coloration toward the top of the head. Pupil circular. Iris turquoise green. Mental region cream to the level of second pair of postmentals grading into salmon, decorated with 1–2 scales thick oblique irregular dark-pigmented stripes directed posterolaterally. Throat dark bronze with touches of salmon, dark pigmentation coming from mental region forms a semireticulate pattern posteriorly, grading into a dark gular background that retains a bronze brightness in preservative.
Neck, dorsal body, and flanks light brown with discrete bronze brightness, decorated with pale salmon spots 1–3 scales in size, and sparser, randomly distributed smaller dark spots, creating a side-to-side coarsely aligned light dotted pattern. Nuchal collar positioned at the level of gular fold, complete ventrally, incomplete dorsally, formed by 3–4 rows of dark scales extending dorsally to the uppermost limit of the flank, outlined by one row of cream scales anteriorly and two rows posteriorly. Uppermost limits of flanks with artichoke green brightness along the second half of the body, toward the tail. Chest pigmented; irregular discontinuous dark stripe positioned before the insertion of forearms, separated from nuchal collar by cream stripe that outlines it posteriorly. Ventral ground coloration grayish cream with sparse light brown pigmentation that faints toward the belly. Forearms light brown; small irregular dark
Males Females
0.6
0.4
0.4
0.2 0.2
1 1
PC 0 PC
0
-0.2
-0.4
-0.2
-0.4
T
.
hygomi sertanejo
T
.
T.
cocorobensismucujensis
T.
T erythrocephalusinsula . nus T. ethe itambereridgei T. T. monta T. torquatusnus T oreadicus . T chromatops .
.
psammonastes T hispidus . T imbituba . T. catalanensis T. T hygomi . mucujensis sertanejo . T T. cocorobensis T erythrocephalusinsula . nus T ethe . itambereridgei T. T. torquatus T. monta T chromatops .
.
psammonastesnus T. oreadicus T hispidus . T. catalanensis T imbituba .
T. T T. T
spots randomly spread over. Anterior plane of thighs light brown, posterior plane and shanks artichoke green; small irregular dark spots randomly distributed. Supracarpal and supratarsal regions light brownish bronze; digits light brown with touches of cream. Black femoral and precloacal patches 22/21 and 16 scales long, respectively. Tail artichoke green grading into light brown toward the tip, ventral side pale cream.
COLORATION IN PRESERVATIVE: Intense bronze coloration on the dorsal head faded. Head and dorsal body brown. Oberhautchen on most dorsal and lateral scales lost, and with it the dotted pattern of the holotype (dorsal pattern was preserved in the allotype, with pale cream spots contrasting against the darker background; see fig. 6). Pale longitudinal stripe below the eyes extends from labials to preauricular fringe, and narrow dark interruptions formed by pigmented vertical bars are more evident against the cream background. Mental region cream. Postmental salmon background absent, dark pigmentation toward the throat preserved, without the original bronze brightness. Nuchal collar well marked, but the row of pale salmon scales outlining it became pale cream. Chest coloration persisted nearly unchanged. Ventral coloration preserved the grayish cream coloration, but ventral surfaces of forelimbs, hind limbs and tail became slightly more pale cream. Artichoke green coloration of the posterior plane of thighs and on shanks became brown, similar to dorsum. Artichoke green brightness along the second half of the body toward the tail absent.
MORPHOMETRICS: Tropidurus sertanejo , n. sp., is one of the four smallest species in the T. torquatus group, with adult males ranging from 60.09–79.92 mm SVL and females from 61.98– 72.46 mm SVL. Reduced mean body size is also observed in T. hygomi , T. mucujensis , and T. cocorobensis (fig. 9; tables 3–4). Because PC1 held the majority of the explained variance and
Males N SVL TL HH EOS HW AL FAL HDL THL SL FOL T. catalanensis 9 (5) 105.71 ± 156.90 ± 15.39 ± 26.21 ± 21.75 ± 18.51 ± 16.34 ± 19.92 ± 23.22 ± 26.12 ± 34.58 ± 11.91 16.69 2.04 2.86 2.72 2.37 2.43 2.59 2.95 3.64 3.07 (85.53– (138.00– (12.22– (21.93– (17.27– (14.37– (12.76– (14.37– (18.21– (22.05– (30.81– 121.79) 183.00) 17.72) 30.17) 25.07) 21.31) 21.48) 22.67) 26.71) 31.48) 38.75) T. chromatops 2 (1) 91.14 ± 140.97 12.87 ± 21.63 ± 19.04 ± 15.21 ± 13.04 ± 15.20 ± 20.87 ± 18.29 ± 27.65 ± 1.73 1.02 1.41 1.21 1.96 0.93 0.12 0.49 0.84 1.03 (89.91– (12.15– (20.63– (18.18– (13.82– (12.38– (15.11– (20.52– (17.69– (26.92– 92.36) 13.59) 22.63) 19.89) 16.59) 13.70) 15.28) 21.21) 18.88) 28.37) T. cocorobensis 11 69.83 ± 102.65 ± 9.96 ± 17.05 ± 13.51 ± 12.42 ± 10.53 ± 13.21 ± 15.09 ± 18.04 ± 25.01 ± (7) 4.75 6.91 1.20 1.33 1.20 1.27 0.74 1.16 1.15 2.46 1.14 (61.39– (94.44– (8.26– (15.37– (11.69– (10.77– (9.31– (10.99– (13.46– (13.56– (23.35– 74.59) 113.00) 12.14) 19.46) 15.84) 14.74) 11.91) 14.59) 17.45) 22.44) 26.92) T. sertanejo , n. sp. 5 (3) 70.24 ± 112.83 ± 9.99 ± 17.83 ± 14.30 ± 12.08 ± 10.72 ± 11.68 ± 14.81 ± 15.95 ± 20.70 ± 9.03 6.54 1.28 2.63 2.47 1.98 1.96 1.08 2.35 4.08 2.38 (60.09– (105.48– (8.44– (14.54– (11.00– (9.85– (8.61– (10.70– (12.86– (10.91– (18.62– 79.92) 118.00) 11.19) 20.03) 16.55) 14.17) 12.72) 13.45) 18.47) 19.94) 24.72) T. erythrocephalus 6 (3) 76.22 ± 120.25 ± 10.52 ± 19.10 ± 15.64 ± 12.52 ± 11.44 ± 13.66 ± 16.60 ± 18.42 ± 24.17 ± 5.97 19.23 0.67 0.80 1.33 1.05 0.66 1.57 1.26 1.08 2.18 (67.90– (99.00– (9.20– (18.07– (13.34– (10.95– (10.55– (11.94– (15.05– (17.23– (21.59– 84.20) 136.46) 10.99) 20.15) 17.46) 13.66) 12.48) 16.04) 18.49) 19.55) 27.85) T. etheridgei 17 76.98 ± 117.91 ± 11.36 ± 19.46 ± 16.02 ± 12.86 ± 11.45 ± 13.78 ± 16.91 ± 18.16 ± 24.02 ± (8) 9.18 8.00 1.57 2.18 2.37 1.68 1.17 1.17 2.24 1.96 1.80 (59.23– (103.79– (8.58– (15.31– (12.00– (9.00– (9.48– (10.50– (13.77– (14.46– (20.23– 96.48) 126.31) 14.37) 23.71) 20.23) 15.89) 13.32) 15.37) 22.27) 21.18) 27.55) T. hispidus 6 (2) 103.30 ± 159.00 ± 14.52 ± 25.91 ± 21.51 ± 17.86 ± 16.65 ± 18.58 ± 22.34 ± 26.84 ± 30.98 ± 10.58 28.28 1.70 2.83 2.53 1.62 1.92 1.84 2.16 3.61 2.67 (84.82– (139.00– (12.01– (20.85– (16.78– (14.94– (13.45– (15.27– (18.65– (20.55– (26.35– 104.82) 179.00) 16.96) 28.47) 23.71) 19.31 18.88) 20.77) 24.74) 30.09) 34.43) T. hygomi 14 62.36 ± 101.72 ± 10.49 ± 16.85 ± 13.45 ± 10.73 ± 9.93 ± 13.18 ± 14.47 ± 18.09 ± 25.86 ± (5) 4.02 8.48 1.19 1.11 1.06 1.41 0.81 0.97 1.48 2.61 1.99 Females N SVL TL HH EOS HW AL FAL HDL THL SL FOL T. catalanensis 16 (8) 80.04 ± 116.76 ± 9.88 ± 18.62 ± 14.95 ± 13.27 ± 11.27 ± 15.45 ± 16.75 ± 18.45 ± 26.15 ± 10.91 16.37 1.47 2.45 2.21 1.99 1.43 1.89 2.39 2.88 2.68 (61.23– (93.00– (7.43– (14.70– (11.50– (9.88– (8.97– (12.57– (13.34– (15.12– (22.19– 102.18) 135.00) 12.63) 23.62) 19.11) 17.34) 14.22) 19.37) 21.98) 25.06) 31.33) T. chromatops 2 (1) 75.20 ± 80.19 11.07 ± 18.37 ± 14.82 ± 12.71 ± 10.87 ± 12.68 ± 16.45 ± 16.85 ± 22.20 ± 3.66 0.21 0.16 0.07 2.42 0.74 0.47 0.93 2.90 0.91 (72.61– (10.92– (18.26– (14.77– (11.00– (10.34– (12.34– (15.79– (14.80– (21.55– 77.79) 11.22) 18.48) 14.87) 14.42) 11.39) 13.01) 17.11) 18.90) 22.84) T. cocorobensis 4 (4) 62.66 ± 85.25 ± 7.97 ± 14.57 ± 11.32 ± 11.21 ± 9.09 ± 12.13 ± 12.40 ± 15.84 ± 21.34 ± 2.33 4.99 0.50 0.70 0.69 1.10 0.64 1.19 1.05 0.64 0.78 (60.27– (78.00– (7.45– (13.99– (10.30– (9.74– (8.18– (10.54– (10.89– (14.89– (20.71– 65.37) 89.00) 8.42) 15.56) 11.80) 12.36) 9.68) 13.07) 13.27) 16.28) 22.47) T. sertanejo , n. sp. 3 (2) 63.37 ± 79.74 ± 8.88 ± 15.69 ± 12.49 ± 10.20 ± 8.64 ± 10.61 ± 12.97 ± 13.13 ± 17.76 ± 1.72 4.00 0.12 0.83 0.41 0.43 0.32 0.45 1.32 2.21 0.43 (61.98– (76.91– (8.74– (15.19– (12.05– (9.70– (8.44– (10.16– (11.45– (11.34– (17.32– 65.29) 82.57) 8.98) 16.64) 12.85) 10.49) 9.00) 11.06) 13.85) 15.60) 18.17) T. erythrocephalus 3 (2) 64.95 ± 88.50 ± 8.38 ± 15.56 ± 12.24 ± 10.36 ± 9.31 ± 11.59 ± 13.99 ± 15.43 ± 19.25 ± 0.91 3.54 0.30 0.25 0.27 0.37 0.24 0.58 0.53 0.52 0.40 (64.08– (86.00– (8.05– (15.28– (11.96– (10.13– (9.07– (10.92– (13.62– (14.83– (18.87– 65.90) 91.00) 8.65) 15.77) 12.50) 10.79) 9.55) 11.93) 14.60) 15.79) 19.66) T. etheridgei 13 (6) 67.66 ± 92.19 ± 9.42 ± 16.26 ± 13.15 ± 11.35 ± 10.14 ± 12.52 ± 13.99 ± 15.75 ± 21.47 ± 7.57 6.27 1.07 1.45 1.95 1.07 0.75 1.30 1.64 1.60 1.33 (59.54– (83.50– (8.04– (13.83– (10.70– (9.84– (9.10– (10.38– (11.71– (13.19– (19.60– 86.28) 103.00) 11.41) 19.22) 17.40) 12.72) 11.94) 14.77) 16.26) 18.67) 23.45) T. hispidus 9 (5) 76.10 ± 113.00 ± 9.73 ± 18.20 ± 14.92 ± 12.67 ± 12.05 ± 14.22 ± 16.13 ± 18.80 ± 23.76 ± 11.18 10.05 1.15 2.16 1.77 1.92 1.81 1.74 2.38 2.46 2.54 (59.83– (98.00– (8.12– (15.33– (12.25– (9.75– (9.52– (11.90– (12.81– (15.15– (20.18– 92.62) 125.00) 11.19) 21.88) 17.86) 15.66) 14.82) 16.44) 19.90) 21.75) 26.93) T. hygomi 10 (6) 52.85 ± 81.11 ± 7.77 ± 13.64 ± 10.43 ± 8.73 ± 8.06 ± 10.53 ± 12.19 ± 15.35 ± 21.11 ± 4.08 13.72 0.65 0.79 0.72 0.68 0.52 0.61 0.88 2.65 0.84
showed positive weights on all variables, it was interpreted as a general measure of size. Although size variation is nearly continuous among Tropidurus species (fig. 9), ANOVA showed a significant difference in mean body size (males: df = 15, sum sq = 6.45, mean sq = 0.43, F value = 23.31, p <0.001; females: df = 15, sum sq = 2.52, mean sq = 0.17, F value = 10.05, p <0.001). Tropidurus sertanejo , n. sp., differed statistically from larger species; pairwise comparisons indicated that males of the new species differ from T. catalanensis , T. hispidus , T. imbituba , T. oreadicus , T. psammonastes , and T. torquatus , while females differ from T. catalanensis , T. hispidus , and T. imbituba (mean body size comparisons for all species pairs shown in appendix 3).
Although MANOVA has identified statistical differences among species centroids (males: df = 15, Wilks’ λ = 0.0022, approx. F = 5.7780, df nun = 165, df den = 977.2494, p <0.001; females: df = 15, Wilks’ λ = 0.0031, approx. F = 4.0070, df nun = 165, df den = 727.0808, p <0.001), PCA plots showed large morphometric overlap for all species analyzed (fig. 10; table 5). The high morphological similarity was statistically confirmed by the low discrimination power of the LDA. The assignment of individuals to species using the LDA function showed 77.44% and 86.67% success rate based on males and females, respectively. These values dropped to 57.14% and 51.43% when the leave-one-out cross-validation procedure was implemented (appendices 4–5). Size-free LDA showed even lower reclassification rates, with 64.6% and 76.19% correct assignment of individuals. The cross-validation procedure resulted in 42.11% and 46.67% correct reclassifications (appendices 6–7).
These results indicate that the morphometric profile of the T. torquatus species group is highly conserved, and body size is an important variable promoting discrimination among species (table 5). For T. sertanejo , n. sp., elimination of isometric size resulted in significant decrease in reclassification success from 60% to 20% in the cross-validation LDA for males and 33% to 0% for females. Males of T. sertanejo , n. sp., were frequently misclassified as T. etheridgei , T. hispidus , or T. imbituba , and females as T. etheridgei , T. erythrocephalus , or T. insulanus (appendices 6–7). The size-free LDA for both sexes indicated that FOL is the most important attribute explaining differences in shape among species. However, the plots of FOL against SVL (not shown) suggest a common allometric trend for all Tropidurus species , except for T. hygomi and, to lesser degree, T. cocorobensis , which have feet proportionally larger with respect to body size (tables 3–4). Therefore, differences in the proportions of FOL in relation to SVL can only be used to discriminate T. sertanejo , n. sp., from two of the four smallest species of the T. torquatus group. Foot length in T. sertanejo , n. sp., corresponded to 26.41%–32.18% of snout-vent length in males and 26.53%–29.32% in females, while values between 32.51%–40.82% and 32.49%–34.56%, and 35.11%–48.42% and 36.01%–44.30%, were observed for males and females of T. cocorobensis and T. hygomi , respectively. Body size and foot length were useful variables distinguishing T. sertanejo , n. sp., from the largest and smallest species of the T. torquatus group, respectively, but morphometric attributes alone were insufficient to separate it from middle-sized forms of the group. Additional morphological attributes are needed to unambiguously distinguish T. sertanejo , n. sp., from most congeners; see “Comparison with other species.”
MERISTICS: All meristic variables showed continuous variation and large overlap among species (fig. 11; table 6). Tropidurus sertanejo , n. sp., had intermediate scale counts and overlapped all other species considerably in number of dorsals, ventrals, gulars, and midbody scales. The
Males Females 0.2 0.2 0.1 2 0.1 2 T. catalanensis PC 0 PC 0 T. chromatops 0.1 0.1 T. cocorobensis T. erythrocephalus 0.2 0.2 T. etheridgei T. hispidus -0.4 -0.2 0 0.2 0.4 0.6 -0.4 -0.2 0 0.2 0.4 T. hygomi PC 1 PC1 T. imbituba T. insulanus T. itambere 2 2 T. montanus 2 ● T. mucujensis 20 LD LD 0 T T.. oreadicus psammonastes -2 -2 T. torquatus ● T. sertanejo , n. sp. -4 -4 -6 -4 -20246 -6 -4 -20246 LD1 LD1
number of tibials and subdigital lamellae on fourth toe were the only variables useful for separating T. sertanejo , n. sp., from other congeners. However, T. cocorobensis , T. imbituba , and T. psammonastes were the only species whose number of tibials or subdigital lamellae was nearly or totally nonoverlapping with T. sertanejo , n. sp. (fig. 11; table 6). Nonparametric MANOVA detected statistical differences in scale counts among species and between sexes, but most of the variance was explained by the former factor (table 7). The absence of interaction between species and sex allowed us to pool individuals of both sexes for the subsequent LDA. LDA confirmed that scale counts are highly similar within the T. torquatus group and meristic variables are not effective in separating most species. The overall reclassification success of the LDA function was 68.84%, and this value dropped to 62.61% when applying the leave-one-out cross-validation procedure. LD1 and LD2 showed large overlap among T. sertanejo , n. sp., and most other species, except for T. catalanensis , T. cocorobensis , T. hygomi , T. imbituba , T. psammonastes , and T. torquatus (fig. 12; table 8). The LDA function had an overall reclassification success of 42.86% for specimens of T. sertanejo , n. sp., and this value dropped to 38.10% when adopting the leave-oneout cross-validation procedure (appendix 8).
The number of subdigitial lamellae on fourth toe had the strongest contribution on both LD1 and LD2, yet not allowing complete separation of any species in both axes. By plotting LD1 against LD2 we were able to identify two species groups that differed in number of scales. The first group included the psammophilous species T. cocorobensis , T. hygomi , and T.psammonastes , characterized by a higher number of subdigital lamellae in relation to the number of dorsals and ventrals. The second included all other species with a relatively lower number of subdigital lamellae. We observed that species with proportionally larger feet had higher numbers of subdigital lamellae on the fourth toe (Pearson correlation: males: r = 0.634, p = 0.008;
TIBIALS GULARS DORSALS
15 20 25 30 60 70 80 90 60 70 80 90 100 110 120 Tita Thyg This This This Tpsa Tser Tpsa Tore Tins Tore Tins Tore Tcoc Tita Tpsa Tery Tser Teth Tita Tery Tchr Tins Thyg Tmuc Tser Tcoc Tery Teth Teth Tmon Ttor Tmon Thyg Tmon Tmuc Ttor Tchr Tchr Tcat Tmuc Timb Tcoc Timb Ttor Timb Tcat Tcat
SUBDIGITAL LAMELLAE MIDBODY VENTRALS
15 20 25 30 35 40 45 50 55 60 60 80 100 120 Tita Thyg This
Tser This Tita
Teth Tcoc Tore
Tore Tery Tins
Tins Tpsa Thyg
This Tore Tpsa
Tery Teth Tser
Tmon Tins Tery
Tmuc Tchr Teth
Tchr Tmuc Tcoc
Ttor Tser Tmuc
Timb Tita Tmon
Tcat Tmon Tchr
Thyg Ttor Timb
Tpsa Timb Ttor
Tcoc Tcat Tcat
Subdigital Species Sex N Dorsals Ventrals Gulars Midbody Tibials
lamellae T. catalanensis All 80 111.99 ± 8.59 80.04 ± 5.36 50.90 ± 4.01 105.44 ± 8.33 27.50 ± 1.81 26.15 ± 1.88
(92–132) (71–97) (41–60) (89–126) (23–32) (22–30)
M 13 105.38 ± 4.96 78.62 ± 3.91 51.15 ± 3.21 97.38 ± 6.53 27.62 ± 2.14 26.38 ± 2.10
(94–110) (71–87) (46–59) (89–106) (24–32) (22–30)
F 47 113.21 ± 8.48 80.91 ± 5.91 50.91 ± 4.56 107.34 ± 7.14 27.43 ± 1.92 25.89 ± 1.91
(96–132) (71–97) (41–60) (97–126) (23–31) (22–30) T. chromatops All 4 101.25 ± 3.86 75.00 ± 2.58 44.75 ± 2.50 92.25 ± 3.59 24.50 ± 1.29 23.00 ± 2.45
(97–105) (72–78) (42–48) (87–95) (23–26) (20–26)
M 2 101 ± 5.66 75.00 ± 4.24 45.00 ± 4.24 91.00 ± 5.66 24.50 ± 2.12 24.50 ± 2.12
(97–105) (72–78) (42–48) (87–95) (23–26) (23–26)
F 2 101.5 ± 3.54 75.00 ± 1.41 44.50 ± 0.71 93.50 ± 0.71 24.50 ± 0.71 21.50 ± 2.12
(99–104) (74–76) (44–45) (93–94) (24–25) (20–23) T. cocorobensis All 16 91.06 ± 5.48 67.31 ± 3.77 39.50 ± 2.42 84.50 ± 4.23 28.56 ± 1.90 27.88 ± 2.13
(84–101) (61–74) (35–43) (78–92) (25–31) (22–31)
M 10 88.3 ± 3.47 66.70 ± 2.95 39.20 ± 2.3 82.20 ± 3.01 28.60 ± 1.78 28.40 ± 1.26
(84–95) (61–71) (36–43) (78–86) (26–31) (27–30)
F 6 95.67 ± 5.28 68.33 ± 5.01 40.00 ± 2.76 88.33 ± 3.01 28.5 ± 2.26 27.00 ± 3.03
(87–101) (61–74) (35–42) (84–92) (25–31) (22–31) T. sertanejo , n. sp. All 21 86.48 ± 7.78 71.10 ± 5.81 45.33 ± 3.4 78.38 ± 6.82 21.90 ± 1.37 20.48 ± 1.72
(73–103) (60–80) (40–53) (65–89) (20–25) (18–23)
M 5 80.60 ± 4.28 68.60 ± 6.31 43.20 ± 2.59 76.00 ± 3.94 22.00 ± 1.41 20.00 ± 1.22
(75-84) (62–76) (40–46) (72–82) (21–24) (19–22)
F 3 93.33 ± 7.37 73.00 ± 11.27 46.67 ± 3.06 84.67 ± 6.66 23.33 ± 1.53 21.67 ± 2.31
(85–99) (60–80) (44–50) (77–89) (22–25) (19–23) T. erythrocephalus All 15 87.07 ± 4.64 67.67 ± 4.92 39.80 ± 3.14 81.47 ± 6.32 25.60 ± 1.96 21.80 ± 0.56
(80–96) (62–80) (35–46) (66–93) (22–29) (21–23)
M 6 83.67 ± 2.42 67.67 ± 3.93 39.33 ± 3.83 76.17 ± 5.64 25.67 ± 2.16 22.00 ± 0.63
(80–86) (62–73) (35–46) (66–80) (23–29) (21–23)
F 6 90.00 ± 2.97 66.33 ± 4.23 40.50 ± 2.81 85.50 ± 3.94 25.83 ± 1.72 21.67 ± 0.52
(86–95) (62–74) (37–45) (82–93) (24–28) (21–22) T. etheridgei All 19 90.84 ± 5.70 71.63 ± 4.78 43.47 ± 3.81 88.58 ± 6.28 24.05 ± 2.27 20.53 ± 1.43
(81–105) (64–79) (36–48) (80–103) (19–28) (18–24)
M 8 90.12 ± 4.85 73.62 ± 5.07 44.62 ± 3.38 87.88 ± 5.57 24.00 ± 1.07 20.88 ± 1.36
(85–100) (67–79) (38–48) (81–100) (23–26) (19–23)
F 7 92.71 ± 7.78 71.57 ± 3.60 43.14 ± 5.11 89.86 ± 8.47 24.86 ± 2.79 20.57 ± 1.62
(81–105) (67–77) (36–48) (80–103) (20–28) (19–24) T. hispidus All 20 67.95 ± 6.61 61.85 ± 4.70 38.85 ± 2.85 65.80 ± 7.68 20.30 ± 1.81 21.75 ± 1.48
(57–88) (54–73) (35–46) (57–90) (16–23) (19–25)
M 7 66.57 ± 5.38 60.71 ± 2.81 37.29 ± 1.80 61.86 ± 3.34 20.14 ± 1.77 22.00 ± 1.91
(57–72) (57–66) (35–40) (57–66) (18–23) (19–25)
F 10 69.20 ± 7.90 62.60 ± 6.19 40.30 ± 3.13 70.10 ± 8.63 20.30 ± 1.95 21.50 ± 1.18
(60–88) (54–73) (36–46) (62–90) (16–23) (19–23) T. hygomi All 23 80.78 ± 4.88 59.74 ± 3.60 38.39 ± 3.07 83.00 ± 5.61 27.35 ± 3.32 27.17 ± 1.77
(72–90) (52–66) (33–46) (72–96) (21–34) (23–30)
M 11 81.27 ± 4.71 60.91 ± 3.21 38.73 ± 3.20 79.00 ± 3.77 28.55 ± 3.42 27.64 ± 2.06
(72–88) (54–66) (34–46) (72–84) (23–34) (23–30)
F 8 81.38 ± 5.63 58.75 ± 3.81 38.00 ± 3.78 88.00 ± 4.84 26.88 ± 3.31 27.25 ± 1.39
(75–90) (52–63) (33–45) (81–96) (21–30) (26–30)
females: r = 0.509, p = 0.044). Positive associations between body size and other meristic variables were similarly expected, but we failed to find significant correlations between SVL and number of dorsal, ventral, gular, and midbody scales. These results are in agreement with our previous observations that SVL and scales counts lack consistent correlation within species (results not shown), suggesting that the number of scales is determined from birth and shows no significant ontogenetic, static, or interspecific allometric trends. Although meristic variables are commonly used to distinguish lizard species, our results showed that we cannot rely on scale counts to separate T. sertanejo , n. sp., from most species of the T. torquatus group.
ETYMOLOGY: The species name, sertanejo , is to be treated as an indeclinable word. In Portuguese, sertanejo is an adjective that alludes to interior or rural areas. It is also used as a noun to refer to the people born in the sertão. In current times, the term sertão is mostly used to refer to the dry areas of northeastern Brazil dominated by the Caatinga biome, where T. sertanejo , n. sp., occurs. Naming T. sertanejo , n. sp., we aim to honor all men and women who bravely thrive in the historically neglected semiarid Brazilian Caatinga.
GEOGRAPHIC DISTRIBUTION AND CONSERVATION STATUS: Tropidurus sertanejo , n. sp., is endemic to the semiarid Brazilian Caatinga and known exclusively from two localities in the State of Bahia, northeastern Brazil. The first is the RPPN Fazenda Pé da Serra, Serra do Arame, Municipality of Ibotirama, a private natural reserve located in the western portion of the state. The second is the Municipality of Caetité, approximately 150 km south of Ibotirama (fig. 3). Although Tropidurus sertanejo , n. sp., has part of its distribution covered by a small protected area (total area of the RPPN Fazenda Pé da Serra: 12.59 km 2; Portaria IBAMA 60-92/N), the fact that only one additional (disjunct) population is known from Caetité represents a critical conservation issue. However, since actual distribution limits, population size, and local abundance of T. sertanejo , n. sp., are unknown, the data currently available allow us to recommend its classification only as “data deficient” according to the criteria proposed by IUCN (2001). In addition to confirming the unique identity of the Caatinga herpetofauna (Rodrigues, 2003; Rodrigues et al., 2003; Carvalho et al., 2013; Guedes et al., 2014), the discovery of T. sertanejo , n. sp., calls attention to the need of extensive mapping of lizard populations within the biome. It also corroborates the existence of higher taxonomic diversity in Tropidurus (Carvalho, 2013) , and makes clear that besides cryptic species even easily diagnosable taxa still remain to be described in this group (A.L.G. Carvalho, unpublished data; M.A. Sena, unpublished data).
ECOLOGICAL REMARKS: At the RPPN Fazenda Pé da Serra, Municipality of Ibotirama, T. sertanejo , n. sp., occurs in syntopy with T. pinima and T. hispidus , although these species exhibit distinct spatial niches and have not been observed using the same microhabitats. With a common behavior found among species of the T. semitaeniatus group, T. pinima was observed using rock crevices in large rock outcrops. Several individuals, including up to five juveniles, were seen basking on the same rock surfaces. Tropidurus hispidus was observed exclusively around an artificial dam inside the RPPN Fazenda Pé da Serra, and seemed limited to rocks and concrete structures in the area. The distribution of this species in the region is apparently limited to human-modified areas, and it remains unclear whether its occurrence is opportunistic or the species is established in localities apart from those sampled during our visits to the reserve. Several adult males and females were observed basking on the walls of the dam, and hiding in cracks or underneath concrete structures during the hottest periods of the day. A few meters south of the main river that crosses the RPPN Fazenda Pé da Serra, we observed T. sertanejo , n. sp., using small to middle-sized rocks (~ 40–120 cm) on trails that cut through sandy areas covered with dry forests and caatingas (fig. 2). The local distribution of the new species seems to be determined by the presence of both rocks on sandy soils, rather than each of these elements individually. Tropidurus sertanejo , n. sp., was never observed using branches or tree trunks, but we confirmed it uses holes underneath rocks as shelters. As we approached the lizards, they fled to the surrounding bushes and remained motionless on the leaf litter where their coloration served as camouflage. When threatened, they also fled to their shelters and hid for a few minutes until returning to their original location on the rocks. In general, the lizards returned to the very same rocks even after being approached several times, which suggests that they defend their territories and have small home ranges of a few square meters. Tropidurus sertanejo , n. sp., is diurnal and active individuals were observed until sunset (approximately 6 P. M.). Nothing is known about its diet. The large extension of the natural landscapes suggests that T. sertanejo , n. sp., occurs in contiguous areas along the Serra do Arame. However, additional fieldwork is needed to determine its actual distribution limits and ecological requirements at the type locality. Specimens of T. sertanejo , n. sp., from Caetité were active on rocky outcrops emerging from a wide plateau of white sands covered by dense thickets of low vegetation. Cactaceae , Bromeliaceae , Velloziaceae , and Euphorbiaceae compose the dominant plant cover at the rocky areas. When approached, lizards took refuge in rock crevices among the vegetation. The species is sympatric but never syntopic with a still undescribed species of Eurolophosaurus (see Passoni et al., 2008) and with another relative of the T. torquatus group preliminarily identified as T. aff. etheridgei . The latter two species were never observed on rocks. They occupy exclusively the adjacent sandy soil areas, moving among thickets of vegetation separated by variable extensions of bare sand.

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |