
Phyxioschema spelaeum, Schwendinger, Peter J., 2009
|
publication ID |
https://doi.org/ 10.5281/zenodo.188258 |
|
DOI |
https://doi.org/10.5281/zenodo.6214234 |
|
persistent identifier |
https://treatment.plazi.org/id/03A9B15A-FFBC-7E56-FF06-B765FBA5FD52 |
|
treatment provided by |
Plazi |
|
scientific name |
Phyxioschema spelaeum |
| status |
sp. nov. |
Phyxioschema spelaeum sp. n.
Figures 18–20 View FIGURE 18 A – E View FIGURE 19 A – N View FIGURE 20 A – D , 21B, D View FIGURE 21 A – D , 22D–I View FIGURE 22 A – I
Material: THAILAND (southern region): Krabi Province, Ao Luk District, Sra Yuan Thong Cave, ca 3 km SE of Ao Luk Tai (8°21'40''N, 98°44'55''E), 60 m, male holotype (matured 2.I.2008), 4 male paratypes (matured 28.XI.2007, 14.XII.2007, hatched 10.X.2007 and matured 7.IX.2008, hatched 31.VIII.2007 and matured 18.XII.2008) and 13 female paratypes, 11.VII.2007 (sample TH-07/07). From the same locality, 1 male paratype (matured 9.II.2009) and 3 female paratypes, 1 juvenile, 21.VI.2008 (sample THMA-08/12). All specimens collected by P.J. Schwendinger and deposited in MHNG.
Etymology: Latin: " spelaeum " (from the Greek "spelaion") = cave; noun in apposition.
Diagnosis: Medium-sized species, distinguished from the similar and geographically close P. eripnastes sp. n. by: Males with stronger apical bristles on palpal tarsus and less curved embolic tip; ventral spur of tibia II proximally narrower; metatarsus II with evenly rounded (not angular) retroventral keel and a plane or slightly depressed C-shaped area at distal end of proventral keel; leg tarsi III–IV with spines, I–II without. Females with longer spermathecae, each with distinct proximal boss on its inner (ental) side; lateral receptacles larger; secondary receptacles absent.
Description: MALE (holotype). Colour in alcohol: Body generally light brown. Dorsal side of opisthosoma light grey-brown, no dorsal pattern discernible ( Fig. 18B View FIGURE 18 A – E , see also Fig. 22G View FIGURE 22 A – I showing a male paratype), its ventral side cream-grey; eye mound black; ventral side of prosoma (including legs) cream; anterior zone on labium and prolateral zone on maxillae white; ventral spur on tibia II orange; spinnerets ventrally mottled with white spots.
Body 9.0 long. Carapace 3.7 long, 3.3 wide, oval, almost flat, covered with adpressed hairs interspersed by darker, more erect bristles (some wavy), these longest on carapace margin, in front and on eye mound, and in posterior part of carapace. Eyes on low mound; eye group ( Fig. 19B View FIGURE 19 A – N ) 0.40 long, anterior eye row slightly procurved, 0.77 wide, posterior eye row slightly recurved, 0.77 wide. Eye diameters and interdistances: AME 0.20, ALE 0.25, PME 0.18, PLE 0.21; AME–AME 0.07, AME–ALE 0.03, PME–PME 0.34, PME–PLE 0.02. MOQ 0.33 long, 0.33 wide anteriorly, 0.62 posteriorly. Fovea pit-like, with 2 long, thickened and slightly procurved foveal setae anterior to it.
Chelicerae weak, grooves with 10/13 prolateral teeth and 15/16 median proximal denticles. Maxillae ( Fig. 19C View FIGURE 19 A – N ) 0.9 long, 0.6 wide, with pallid prolateral zone; anterior lobe indistinct, with quite wide but fairly indistinct serrula on ridge. Labium ( Fig. 19C View FIGURE 19 A – N ) 0.3 long, 0.6 wide, anterior edge distinctly setose, followed by pallid zone; posterior part pigmented, with few fine setae. Sternum ( Fig. 19C View FIGURE 19 A – N ) 1.9 long, 1.6 wide, cordate, with deeply excavated post-labial depression formed by fused anterior sigilla and labio-sternal groove, and with 3 pairs of indistinct marginal sigilla.
Palps ( Fig. 19D–E View FIGURE 19 A – N ) set with stiff bristles, including 6/7 thick apical bristles on tarsus (these stouter than in other congeners, but longer, and therefore relatively more slender, than cymbial spines of an Euagrus chisoseus Gertsch male from Texas in MHNG). 6+6 trichobothria in 2 rows on tibia, 8 in a zig-zag row on tarsus. Palpal bulb at base of embolus fairly wide; embolus quite short, mostly straight, with indistinctly curved tip.
Legs 2134. All tarsi pseudosegmented; tarsi III and IV with spines, tarsi I and II aspinose. Preening combs absent. I: tibia cylindrical, with strong prolateral, ventral and retrolateral spines in distal two-thirds but none in proximal third ( Fig. 19G View FIGURE 19 A – N ); patella with a row of 4 sigmoid spines retroventrally, without triangular projection on retrolateral margin ( Fig. 19F View FIGURE 19 A – N ); femur with medium-long, rather narrow band of hooked spinules retrodorsally ( Fig. 18D View FIGURE 18 A – E ). II: metatarsus ventroproximally with 2 keels ( Fig. 19L View FIGURE 19 A – N ), proventral one ( Fig. 19K, L View FIGURE 19 A – N ) sharp, distally ending in C-shaped depression, and situated sligthly more distally than retroventral keel ( Fig. View FIGURE 19 A – N
19L), latter widely rounded and protruding sideward ( Fig. 19L–N View FIGURE 19 A – N ); tibia slightly incrassate, band of elongated spinules on prolateral side straight, narrow, slightly inclined from longitudinal axis of tibia, reaching height of distal side of ventral spur ( Fig. 18C View FIGURE 18 A – E ); ventral spur of tibia with indistinctly bilobed apex carrying a pair of equally long and parallel megaspines ( Fig. 19H View FIGURE 19 A – N ); femur with long band of hooked spinules proventrally ( Fig. 18E View FIGURE 18 A – E ). Spination: I: patella d1, r3; tibia p8/9, r4, v3; metatarsus v4 / 5. II: patella d2; tibia p2, v2 megaspines; metatarsus p2, v 6. III: patella d3; tibia d2, p2, v6, r2; metatarsus d6, v7; tarsus p0/1, r0/3 (3 on left side). IV: patella d3; tibia d1/2, p2, v6, r2; metatarsus d7, v7; tarsus r1. Trichobothria: 2 rows of 6–8 each on tibiae, 10–12 in single row on metatarsi, 10–12 in single row on tarsi. Paired claws with 11–13 teeth in sigmoid row, unpaired claw with 4–6 sessile teeth.
Opisthosoma 4.5 long, 3.3 wide; dorsal side quite densely covered with fine light adpressed hairs, short dark hairs and long strong dark bristles with darkened sockets ( Fig. 18B View FIGURE 18 A – E ); ventral side only with short and medium-sized dark hairs. Posterior median spinnerets 0.6 long; posterior laterals 5.8 long (proximal article 1.7, median article 1.6, pseudosegmented distal article 2.5).
FEMALE ("allotype"). As the male, except for: body generally darker (more orange-coloured); chelicerae, labium, maxillae, sternum, tips of palpal and leg tarsi, booklung covers and anus distinctly darker; pale prolateral zone of maxillae and pale anterior zone of labium more distinct.
Habitus as in Fig. 22D View FIGURE 22 A – I (showing a different female). Body 11.7 long. Carapace 4.0 long, 3.2 wide. Eye group ( Fig. 19A View FIGURE 19 A – N ) 0.39 long, anterior eye row 0.80 wide, posterior eye row 0.87 wide. Eye diameters and interdistances: AME 0.19, ALE 0.24, PME 0.18, PLE 0.21; AME–AME 0.06, AME–ALE 0.03, PME–PME 0.36, PME–PLE 0.02. MOQ 0.36 long, 0.39 wide anteriorly, 0.64 posteriorly.
Chelicerae stronger than in male, grooves with 14 prolateral teeth and 25/28 median proximal denticles. Maxillae 1.2 long, 0.7 wide, serrula wider and more pronounced than in male. Labium 0.3 long, 0.8 wide. Sternum 1.9 long, 1.7 wide.
Palps with 7+7 trichobothria on tibia and 11/12 on tarsus. Tarsal claw with 16/17 teeth.
Legs 2134. All tarsi integral; tarsi II–IV armed with spines. Spination: I: patella d1; tibia p1, v5 (indistinct); metatarsus v7 / 8. II: patella d2; tibia p1, v5 (indistinct); metatarsus p2, v6 /7; tarsus p1, v0/1, r1/ 2. III: patella p2, r1; tibia d2, p2, r2, v6 (indistinct); metatarsus d6/7, v7; tarsus p0/1, r 2. IV: patella p2, r1; tibia p2, d1, r2/3, v6 /7 (indistinct); metatarsus d6/8, v7 /8; tarsus p1, r1/2. Trichobothria: 2 rows of 7–8 each on tibiae, 11–13 in single row on metatarsi, 10–12 in single row on tarsi. Tarsal organ low, with shallow concentric ridges ( Fig. 21B View FIGURE 21 A – D ). Paired claws with 12–16 teeth in sigmoid row, unpaired claw with 4–8 sessile teeth ( Fig. 21D View FIGURE 21 A – D ).
Opisthosoma ( Fig. 18A View FIGURE 18 A – E ) 6.0 long, 4.5 wide. Posterior median spinnerets 0.7 long; posterior lateral spinnerets 6.2 long (proximal article 1.8, median article 1.7, distal article 2.7).
Vulva ( Fig. 20A View FIGURE 20 A – D ) with 2 proximally wide spermathecae, each with distinct proximal boss on inner (ental) side; only 2 receptacles present; lateral receptacle short, with completely sclerotised, slightly constricted neck and moderately large head lacking pores; median receptacle with moderately convoluted stalk and large head lacking pores; secondary receptacles absent.
Palp and leg measurements: See Table 1.
Variation: Measurements of males (n=6) (females with egg sacs in parentheses; n=7): body length 8.6–9.2 (9.8–11.3), carapace length 3.4–3.7 (3.4–4.0), width 3.0–3.3 (2.9–3.5). Most specimens (from a total of 23) possess two foveal setae, one has only one seta, two have three setae. One male paratype has a spine on tarsus II of one side, another lacks spines on tarsus III on one side; other males have spines on tarsi III–IV of both sides. Seven (out of 16) females, including the "allotype", lack spines on tarsus I; six have a spine on one side of tarsus I, one has a spine on both tarsi I; one female has two spines on tarsus I of one side, but none on the other, another female has two on one side and none on the other. Two male paratypes have three retroventral spines on both patellae I, one male has three on one side and four on the other, the other three males (including the holotype) have four patellar spines on both sides. In three males (including the holotype), the two megaspines on the ventral spur of tibia II are slightly convergent, with their tips almost touching ( Fig. 19H View FIGURE 19 A – N ); in another male the tips are overlapping; in two other males the megaspines are parallel to each other ( Fig. 19I View FIGURE 19 A – N ). Of the six vulvae examined, one has a single pore on each median receptacular head ( Fig. 20B View FIGURE 20 A – D ), one has three pores on one median receptacular head ( Fig. 20C View FIGURE 20 A – D ), all others have none; four females have pores on the lateral receptacular head ( Fig. 20B–D View FIGURE 20 A – D ) and two (including the "allotype") lack pores on all receptacular heads ( Fig. 20A View FIGURE 20 A – D ).
Relationships: The shape of the lateral receptacles shows that P. s p e l a e u s sp. n. is most closely related to P. eripnastes sp. n. (lateral receptacular bases short, quite wide and completely sclerotised in these two species) and that both are sister to P. s a y a m e n s e sp. n. and P. huberi sp. n. (lateral receptacular bases completely sclerotised and prolateral band of elongated spinules on tibia II straight in all four species).
Distribution and habitat: This species is known only from the limestone hills east of Ao Luk Tai ( Fig. 1 View FIGURE 1 , locality 18). Most spiders were collected from the twilight zone behind the wide entrance (with a pond) of a cave at the foot of an isolated limestone hill; only few spiders were seen in the completely dark parts of that cave. Webs retreating into holes in rock and stal were found at the base of cave walls and of columns ( Fig. 22E View FIGURE 22 A – I ). A few webs were also seen outside the cave, on large, isolated limestone rocks in a semi-evergreen rain forest about 1 km away, but surprisingly not on the nearby cliffs of the main ridge of limestone hills.
Phenology: ( Table 3): Four males were collected in June and July, and subsequently matured between late November and early February. A fifth male hatched in captivity at the end of August and became mature in late December of the following year. A sixth male hatched in October and became mature in the following September, presumably earlier in the year than males in nature. Males thus have a lifespan of not more than 1.5 years; they continued feeding for only about two weeks after reaching maturity. An egg sac containing newly hatched first instar spiderlings was found in the cave in early June, more such egg sacs in late June. Three females (captured in June and July) constructed egg sacs in captivity between the end of July and late October; spiderlings hatched almost six weeks after oviposition. Two other females mated in captivity in early January; one of them (but not the other) produced an egg sac more than six weeks later, in late February, and again in early April and late May. Egg sacs, suspended in the web outside the retreat and camouflaged with particles of limestone ( Fig. 22F View FIGURE 22 A – I , see small arrows), contained 15–37 light yellow eggs (n=7). Females which built egg sacs in July to October moulted between October and December; one with egg sacs in February to May moulted in September; others moulted in July to March.
The presence of eggs in the cave about four months after the short-lived males became mature suggests that, in nature, inseminated females produce eggs for much longer than females did in captivity. An alternative explanation is that a second mating period in April and/or May was overlooked (see also the phenology of P. eripnastes sp. n.). Inspecting webs in the cave at night, when all spiders were sitting outside their retreats (only a few did so during daytime), in early June and again in late June, however, revealed no adult males.
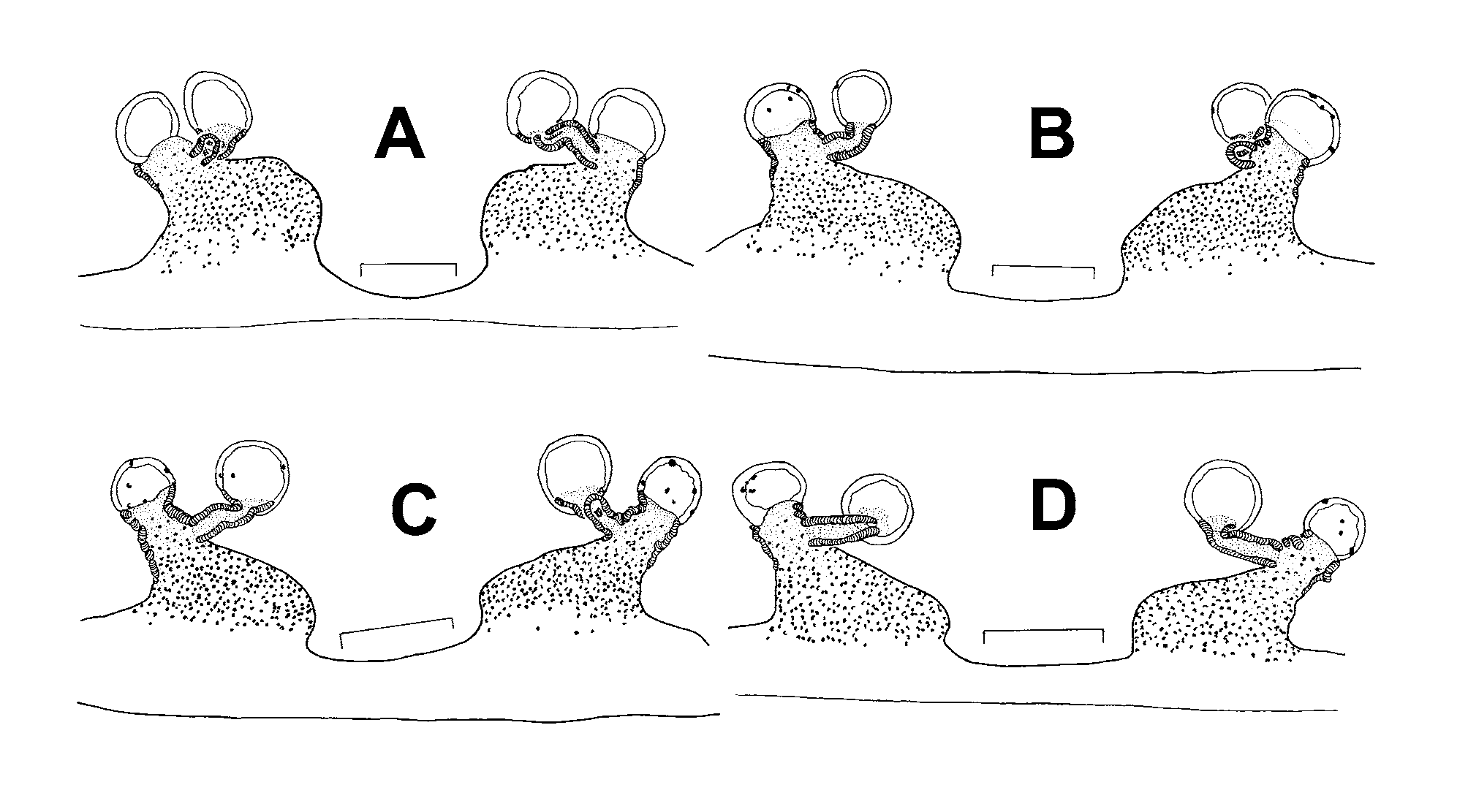
Mating behaviour: Three matings (of different pairs) were observed in January 2008 and 2009. Copulation lasted 35 to 86 minutes during which the male held his opisthosoma bent ventrally by almost 90° so that its tip was in (or in near) contact with the opisthosoma tip of the female. No such mating posture was observed in other Phyxioschema species. During courtship the male produced some light jerk-quivers and drummed on the female with his first pair of legs, but he did not drum on the web with his palps. The female responded by advancing and also drumming on the male with her first pair of legs. Adoption of the mating position was quick and vigorous, as in other Phyxioschema . During interlocking, the female repeatedly became “restless” but was “quieted” again by the male by means of vigorous jerks.





| MHNG |
Museum d'Histoire Naturelle |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |