
Paguristes puniceus Henderson, 1896
|
publication ID |
https://doi.org/10.5281/zenodo.158499 |
|
DOI |
https://doi.org/10.5281/zenodo.5625071 |
|
persistent identifier |
https://treatment.plazi.org/id/03A8F02D-743C-FFCA-D66F-FABA28FAFCD0 |
|
treatment provided by |
Plazi |
|
scientific name |
Paguristes puniceus Henderson, 1896 |
| status |
|
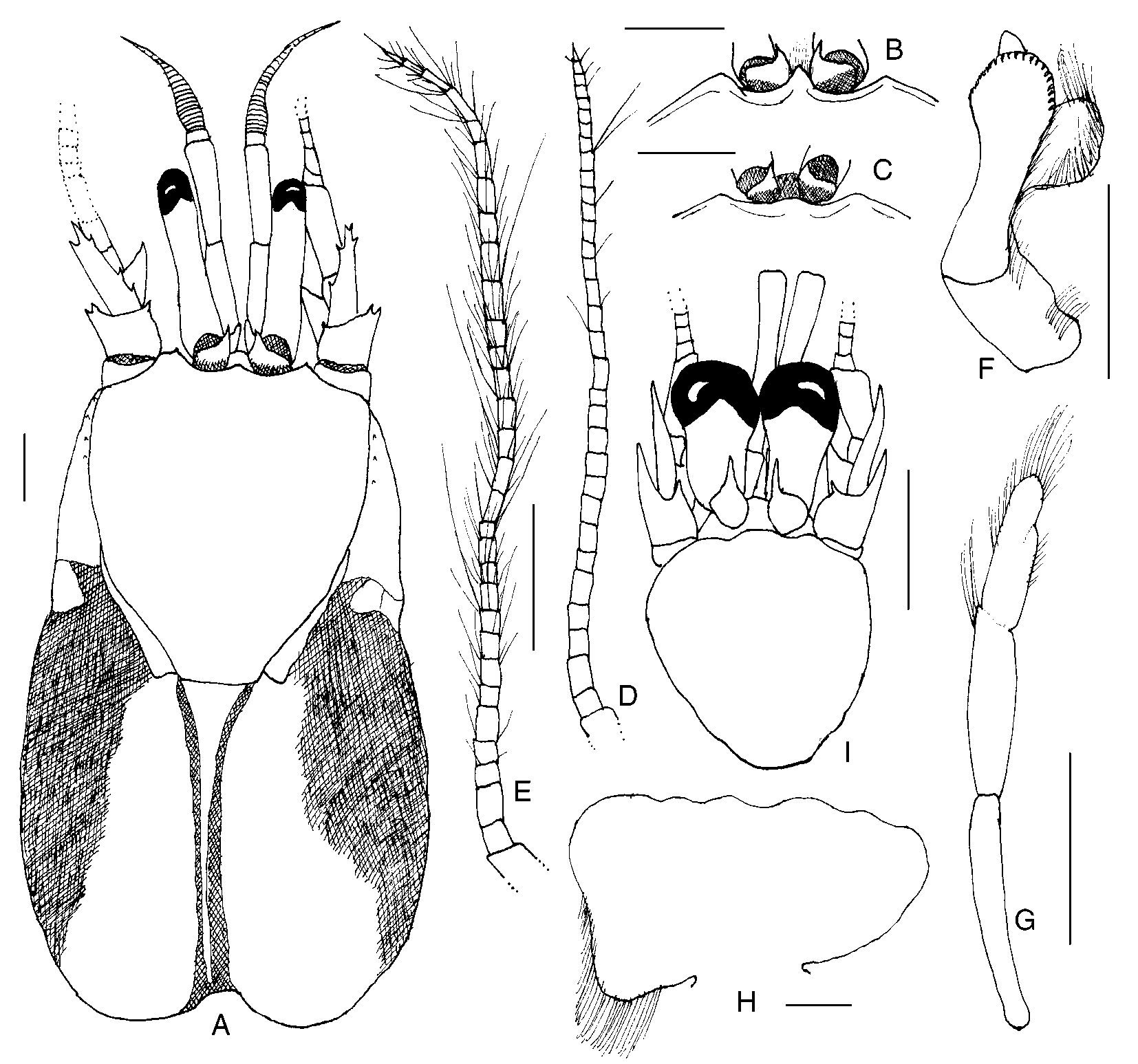
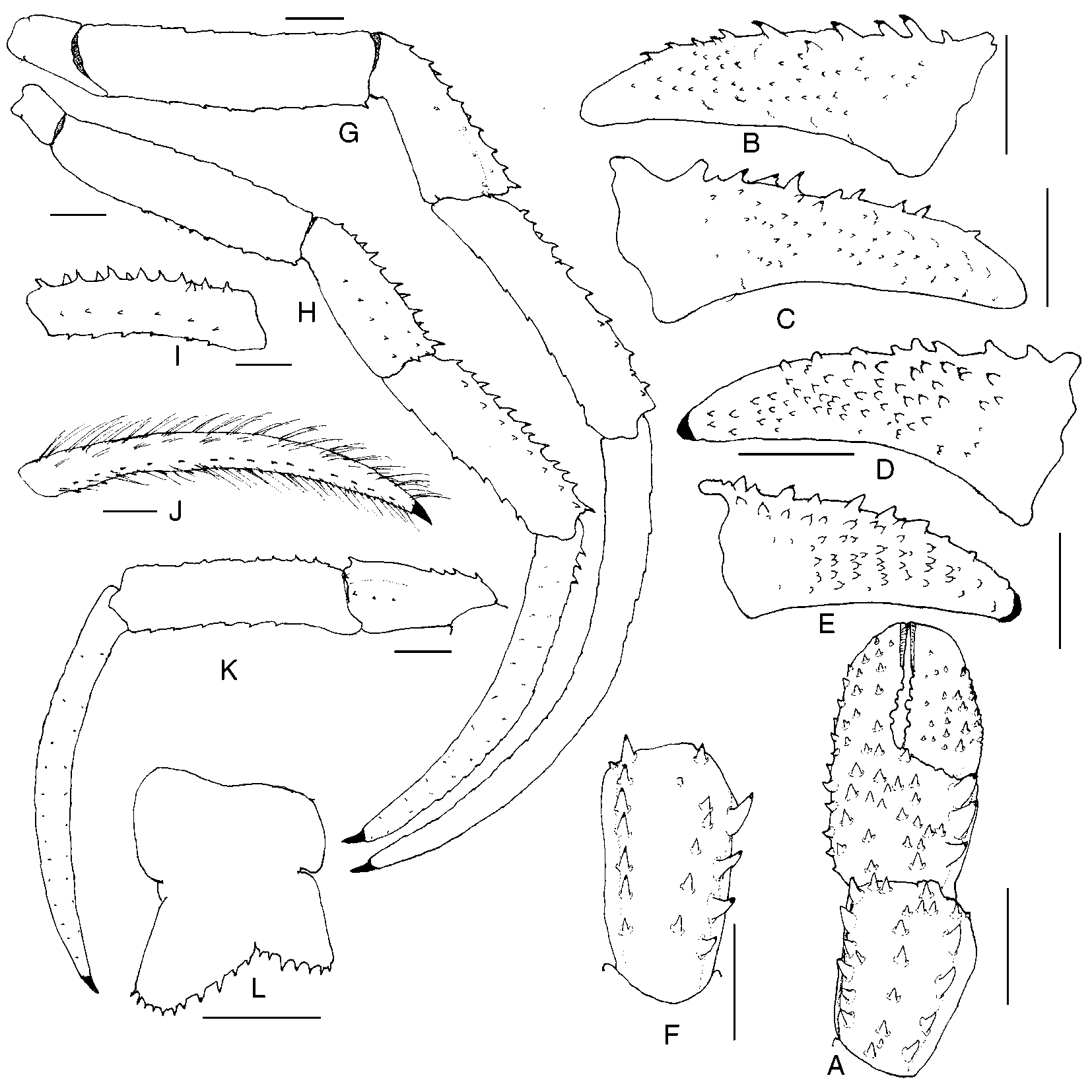
Paguristes puniceus Henderson, 1896 View in CoL ( Figs. 1 View FIGURE 1 , 2 View FIGURE 2 )
Paguristes puniceus Henderson, 1896: 527 View in CoL ; Alcock & Anderson, 1897: pl. 32, fig. 1; Alcock, 1905: 38, pl. 3, fig. 6; Kemp & Sewell, 1912: 25; Balss, 1924: 768; Gordan, 1956: 323 (literature); Komai, 2001: 423.
Paguristes puniceus View in CoL var. unispinosa Balss, 1912: 92 .
Not Paguristes puniceus View in CoL . – Miyake, 1978: 38, fig. 13.
Redescription. Thirteen pairs of biserial gills. Shield ( Fig. 1 View FIGURE 1 A) slightly longer than broad; dorsal surface sometimes with few spines or spinules marginally and/or laterally, and often with tufts of moderately long setae, particularly laterally. Rostrum ( Fig. 1 View FIGURE 1 A–C) narrowly to broadly triangular, rarely broadly rounded and nearly obsolete; not reaching or slightly overreaching level of lateral projections, rarely reaching midlength of ocular acicles, unarmed or with terminal spine or spinule, sometimes also with few long simple setae. Lateral projections ( Fig. 1 View FIGURE 1 A–C) obtusely triangular to subquadrate, sometimes simply broadly rounded, with or without terminal spine or spinule. Branchiostegites often with few widelyspaced spinules or small spines on dorsal margins distally or lateral surfaces dorsally, sometimes with 1 or 2 irregular rows of very small spines; usually with 1–3 small spines to tiny spinules on anterior margins. Posterior carapace with posterior median plate ( Fig. 1 View FIGURE 1 A) extremely narrow to moderately broad, well calcified or not.
Ocular peduncles 0.3–0.7 length of shield; dorsal surfaces each sometimes with row of long setae; corneal diameter 0.1–0.3 of peduncular length. Ocular acicles usually triangular, occasionally subovate, frequently only distal halves calcified; each terminating acutely, subacutely or bluntly, often with terminal spine; separated basally by 0.5 to entire basal width of 1 acicle.
Antennular peduncles when fully extended overreaching distal margins of corneas by 0.1 length of ultimate segment to 0.2 length of penultimate segment. Ultimate segment often with row of very fine setae on dorsal surface; penultimate segment glabrous; basal segment usually with 1 or 2 spinules on distolateral margin, ventrolateral distal margin produced into elongate spiniform process.
Antennal peduncles usually reaching from bases of corneas to distal corneal margins, occasionally not reaching corneal bases or overreaching distal margins of corneas by 0.2 of fifth peduncular segment. Fifth segment sometimes with tuft of setae at dorsolateral distal margin and with few scattered moderately long setae on other surfaces; fourth segment commonly with small dorsodistal spine or spinule; third segment with prominent ventrodistal spine; second segment with dorsolateral distal angle produced, usually terminating in bifid spine, occasionally 1 or 2 smaller spines on lateral margin distally and tufts of long setae, dorsomesial distal angle unarmed or with 1 spine. Antennal acicle reaching from 0.2 of fifth peduncular segment to slightly overreaching distal segmental margin, with prominent terminal, most frequently bifid, spine; mesial margin with 0–6 spines, lateral margin with 0–3 spines, sometimes partially obscured by long setae. Antennal flagellum varying from only as long as shield to slightly longer than entire carapace, each article naked, sometimes with only 1 or 2 long setae ( Fig. 1 View FIGURE 1 D), or frequently with numerous setae, becoming longer and denser distally ( Fig. 1 View FIGURE 1 E).
Chelipeds ( Fig. 2 View FIGURE 2 A) approximately equal or occasionally subequal, left or right slightly larger; armature generally similar; dactyl and fixed finger usually with slender, sometimes moderately broad, hiatus proximally. Dactyl with 13 rows of spines or spinulose tubercles on dorsomesial margin, varying in size from low, subacute tubercles to large, acute, corneoustipped spines, often accompanied by tufts of moderately long setae; dorsal surface unarmed or with few to numerous tubercles, spines or spinules and tufts of setae; mesial face ( Fig. 2 View FIGURE 2 B–E) with irregular, vertical, oblique or longitudinal rows of very small, often corneoustipped spinules or spines, frequently accompanied by short stiff setae; spines and spinules usually fewer in number on right dactyl, decreasing in size and number distally on both dactyls; cutting edge with row of small calcareous teeth in proximal 0.4–0.5, corneous teeth distally; terminating in small corneous claw and frequently slightly overlapped by fixed finger. Palm with row of 3 or 4 moderate to prominent spines and tufts of setae on dorsomesial margin, dorsolateral margin not delimited; weakly convex dorsal surface with 4–6 irregular rows of somewhat smaller spines, each accompanied by tuft of setae, 3 or 4 rows extending nearly entire length of fixed finger; lateral surface with rows of tubercles and tufts of setae; mesial face with few low tubercles and sparse tufts of setae, ventral surface with row of tubercles and tufts of setae extending almost entire length of fixed finger; cutting edge of fixed finger with row of small calcareous teeth; terminating in large corneous claw. Carpus ( Fig. 2 View FIGURE 2 A, F) with row of 4–6 prominent spines on dorsomesial margin, irregular row, often double distally, of smaller tuberculate spines usually mesiad of midline, but occasionally median or laterad, and moderate to smaller row on weakly delimited dorsolateral margin, each accompanied by tuft of setae; lateral face with scattered small tubercles and sparse setae; mesial face with row of tubercles and tufts of setae, 2 to several spinules on distal margin. Merus with 1–3 small to moderately large spines and moderately long setae on dorsodistal margin; dorsal margin with subdistal, transverse row of moderately large to small spines and setae, remaining dorsal margin with row of short spinulose ridges or spinulose protuberances accompanied by tufts of long setae; mesial and lateral faces weakly spinulose, ventromesial and ventrolateral margins each with row of small spines or spinules and long setae, smallest usually laterally. Ischium unarmed or with few spinules on ventromesial margin.
Second ( Fig. 2 View FIGURE 2 G–I) and third ( Fig. 2 View FIGURE 2 J, K) pereopods with dactyls 1.4–2.4 length of propodi; dorsal margins each with row of tufts of moderately long setae, few to short row of small to moderate, often corneoustipped spines proximally always on second pereopods, sometimes also on third; lateral faces each with 2 or 3 longitudinal rows of sparse tufts of short setae; mesial faces each with 2 or 3 rows of tufts of setae and commonly ventral row of very small corneous spines, particularly on third pereopods; row of 14–34 small to tiny corneous spines on each ventral margin often partially concealed by long, stiff setae. Broad dorsal surfaces of propodi of second pereopods each with row of spines and most frequently additional row of smaller spines, spinules, tubercles or protuberances accompanied by tufts of long setae; dorsal surfaces of third with low protuberances and tufts of long setae, and not infrequently also row of smaller spines or spinules; lateral faces each with 1 or 2 rows of tufts of very short setae; mesial faces each with dorsal and ventral row of tufts of setae, frequently associated with protuberances or spinules; ventral margins each with row of spinulose protuberances or tuberculate spines, often corneoustipped, and tufts of setae, spines better developed on second pereopods. Carpi each with tufts of long setae and 2 irregular rows of moderately small to moderately large spines on dorsal margins (second) or small dorsodistal spine and frequently 1 or 2 to rows of smaller additional spines (third); lateral faces each with weak longitudinal sulcus and 2 or 3 rows of tufts of short to moderately long setae, frequently with 1 to several small spines distally, and occasionally median row of spinules; ventral surfaces with few tufts of moderately long setae. Meri each with dorsal row of low protuberances and tufts of long setae; ventral margins each with row of small spines and tufts of moderately long setae (second) or only low protuberances and long setae (third); ventrolateral distal angle with 0–3 spinules (second), or 1 small spine (third) and with tufts of moderately long setae. Ischia of second pereopods each often with few small spinules on ventromesial margin concealed by long setae; third unarmed, but with tufts of long setae. Fourth pereopods each with long, slender preungual process at base of claw; propodi often with moderately dense setation, sometimes more abundant on left; carpi each with or without small dorsodistal spine.
Male first pleopod ( Fig. 1 View FIGURE 1 F) with tuft of setae on outer margin of basal lobe; inferior lamella with row of setae on outer margin, distal margin with 1 row of curved spines extending considerable distance along outer margin; internal lobe moderately small, with row of long setae on inner margin; external lobe often extending slightly beyond distal margin of inferior lamella, occasionally shorter. Second pleopods ( Fig. 1 View FIGURE 1 G) with elongate basal segment, naked or with few setae; endopod with few fine setae on inner margin and tuft of longer setae terminally; appendix masculina with long setae marginally and on inner face. Left pleopods 35 with endopods vestigial or rudimentary. Female paired first pleopods 2segmented. Brood pouch ( Fig. 1 View FIGURE 1 H) varying from subtriangular to subquadrate, sometimes almost fanshaped, margins usually weakly scalloped, with or without marginal long setae. Tergal thickenings above acetabula of pleopods 2–4 sometimes with fringe of long dense setae, setation sometimes restricted to tergal thickening above fourth pleopod, occasionally entirely absent.
Telson ( Fig. 2 View FIGURE 2 L) with moderately deep lateral incisions separating anterior and posterior portions; asymmetrical posterior lobes separated by Vshaped median cleft; left lobe usually appreciably elongate, subtriangular with rounded apex, terminal margin with row of small to moderately small, sometimes widelyspaced spines, usually not continued onto lateral margin, and sometimes partially concealed by accompanying long setae; right lobe with terminal margin rounded, with row of small, often moderately widelyspaced spines, accompanied by long setae, not extending onto lateral margin.
Juvenile development. One juvenile ( Fig. 1 View FIGURE 1 I) male (sl = 2.7 mm) was present among the adults at Investigator station 162. Although it cannot be said with absolute certainty that it is a juvenile of P. puniceus , there is no question that it is a juvenile Paguristes from the development of paired gonopods. However, as the only other species of the genus collected on the cruise, P. pusillus Henderson, 1896 , came from Investigator station 175 off Sri Lanka ( Ceylon) at a depth of 51 m, it seems reasonable that the juvenile from station 162 is referable to P. puniceus . Based on data provided by Provenzano & Rice (1966) for P. sericeus MilneEdwards, 1880 , male gonopods do not develop in Paguristes until after the tenth crab stage. Although the first gonopods in the juvenile were relatively rudimentary, the second were quite elongate, suggesting a considerably later crab stage. However, the rostrum was still represented by a broadly rounded lobe and there was no definitive lateral projection development. Spination was just beginning to develop on the right and only remaining cheliped, and no spines were present on the second or third pereopods. Most interesting were the ocular peduncles and antennal flagella. The ratio of shield length to ocular peduncle length was 0.60, while the ratio of corneal diameter to peduncular length was 0.67; adult ratios in males were 0.50–0.73 for shield to ocular peduncle, but only 0.17–0.24 for corneal diameter to peduncular length. The antennal flagella were more than four times longer than the total carapace length in the juvenile, in contrast the adult male condition of being slightly shorter to slightly longer than the carapace. If this juvenile indeed is P. puniceus , it demonstrates the major morphological changes associated with growth and maturity in the species, which far exceed those reported by Provenzano & Rice for P. sericeus .
Coloration (in life). “Legs and anterior part of carapace light pink. Eyestalks rather darker pink.” ( Henderson, 1896).
Habitat. Reported from substrates of mud and sandy mud; inhabiting gastropod shells, frequently with attached, unidentified Epizoanthus .
Distribution. Bay of Bengal and Travancore coast, India; Andaman Sea; Indonesia; northern Queensland, Australia, 150– 766 m.
Remarks. The Henderson collection of P. puniceus in The Natural History Museum consists of four lots, two from Investigator station 162, one from station 166, and one from station 235, the latter not mentioned in Henderson’s (1896) original description. Of the lots from station 162, one (NHM 96.9.8.22) contains one male and one female with a label indicating “? Syntypes.” The second lot (NHM 1903.4.5.193–200) includes 43 adults and one juvenile with a label reading “Cotypes.” Henderson referred to the specimens from station 162 only as a “large series … ” From Investigator station 166, Henderson reported only one large male. However, the lot from station 166 (NHM 1903.4.5.201–202) presently contains a large male and an ovigerous female. The label reads “compared with type.” It would appear that Henderson, or a subsequent investigator , inadvertently included one of the females from station 162 with the male from station 166. Although the male from station 166 is not catalogued as a syntype, it and the accompanying female are, based on Henderson’s original description and the consecutiveness of the catalogue numbers, considered in this report as part of the syntypic series. The lot from Investigator station 235, collected four years later, also is labeled as having been “compared with the type.”
Henderson’s (1896) description of Paguristes puniceus emphasized the relatively short ocular peduncles and extremely narrow posterior median plate of the carapace. Henderson related P. puniceus to P. setosus (H. Milne Edwards, 1848) , but compared it to Ortmann’s (1892) interpretation of the latter species; however, Paguristes setosus sensu Ortmann was shown by Forest & McLaughlin (1998) not to represent H. Milne Edwards’ (1848) taxon. Henderson differentiated the two taxa not only by posterior median plate configurations, but by differences in ocular peduncle and antennal acicle lengths. Alcock’s (1905) description of P. puniceus included the notation that the ocular acicles were simple or weakly bifid, information presumably gleaned from the specimen(s) he had seen from the Travancore coast that were not part of Henderson’s (1896) original material. It was Alcock’s (1905) remark that Balss (1912) apparently misinterpreted, because in his description of P. puniceus var. unispinosa Balss commented that his taxon agreed with Alcock’s (1905) description except that the ocular acicles were not bifid.
Miyake (1978) reported both P. s e t o s u s sensu Ortmann and P. puniceus from Japanese waters. However, Forest & McLaughlin (1998), recognizing that Ortmann (1892) had incorrectly identified H. Milne Edwards’ (1848) species, reexamined four of Miyake’s (1978) P. setosus and redescribed three of them, together with supplemental material in the collections of the Muséum national d’Histoire naturelle, as P. miyakei . In their remarks, Forest & McLaughlin (1998) differentiated P. m i y a k e i only from H. Milne Edwards’ (1848) P. setosus (as Pagurus ), believing that Miyake’s (1978) P. s e t o s u s represented an incorrect identification by Miyake of Ortmann’s (1892) misinterpreted P. setosus .
As previously indicated, in his reevaluation of Miyake’s species of Paguristes, Komai (2001) was of the opinion that Miyake (1978) had confounded three species under the name P. puniceus . Five of the 11 specimens from Miyake’s collections that Komai (2001) reexamined, he assigned to his new species, Paguristes doederleini Komai, 2001 ; five he believed represented an undescribed species, and one he identified as P. miyakei . Of the latter specimen, Komai said that it closely fit Miyake’s (1978) text figure 39a–d. Obviously, that was a typographical error, because Miyake’s text figure 39 is of Lophopagurus (Australeremus) triserratus ( Ortmann, 1892) (as Pagurus ).
Komai (2001) also reexamined Ortmann’s (1892) specimens identified as P. setosus and concluded that they, as well as the five of Miyake’s (1978) specimens identified as P. puniceus , belonged in the taxon he described as P. doederleini . Komai (2001) distinguished P. doederleini from Henderson’s (1896) P. puniceus by the presence in the former species of a median row of spines on the carpus of the cheliped and fewer setae on the antennal flagellum. Paguristes doederleini was said to be immediately distinguishable from P. miyakei by the shorter antennular peduncles, much more setose mesial faces of the ambulatory propodi and the presence of a ventral row of small corneous spines on the mesial faces of the dactyls of the third pereopods. Komai’s (2001) differential diagnosis of P. puniceus was based on the descriptions that had been given by Henderson (1896) and Alcock (1905), both of which, regrettably, provided minimal pertinent information. Dr. Komai kindly made it possible for the author to compare the syntypic material of P. puniceus with specimens of P. doederleini , P. m i y a k e i, and an undescribed species, albeit not the undescribed taxon confounded by Miyake (1978). That examination has shown that while P. doederleini and P. miyakei are morphologically very closely allied with P. puniceus , both represent species distinct from Henderson’s (1896) taxon. Komai’s undescribed species is also very similar to P. puniceus ; however, differences in ocular peduncle color (white in the Japanese species, pink in P. puniceus ), breadth of the posterior carapace plate, and the absence of spines on the mesial faces of the ambulatory dactyls make conspecificity unlikely (Komai, pers. comm..). In the case of the posterior carapace plate, the specimens from the Andaman Sea and Indonesia that were also generally smaller did not exhibit the marked “funnellike” constriction of the plate seen in the Bay of Bengal specimens. A similar sizerelated condition has been noted in specimens of Paguristes frontalis (H. Milne Edwards, 1836) where the moderately broad plate seen in juveniles becomes increasingly more “funnelshaped” in specimens of larger sizes (personal observation). However, it is doubtful that animal size is the only factor influencing this plate configuration and/or calcification. Habitat and/or shell selection very probably also are contributing factors. The development of spines on the mesial faces of the dactyls may or may not be influenced by animal size. In the type series of P. puniceus , the mesial faces of the dactyls of the third pereopods in individuals occasionally completely lacked spines or only a reduced number of small, weakly chininized spinules were present. In contrast, a row of well developed, prominent, corneous spines was seen on those surfaces in larger specimens.
Henderson (1896) provided no information on coloration for the specimens from station 162, but stated that a note by SurgeonCaptain Anderson accompanying the male from station 166 described pink as the living color of the animal. As previously noted, the coloration of the eyestalks of the undescribed Japanese specimens, as shown in a photograph, are white with only patches of reddishorange distally and proximally. Although pigmentation, as well as morphology, may be variable in P. puniceus , as it is in certain other paguroids (cf. McLaughlin & Provenzano 1974: 181, Haig & Ball 1988: 184), until smaller specimens from the Bay of Bengal become available for comparison, I concur with Dr. Komai’s hesitation to include Japan in the distribution of Henderson’s species.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Paguristes puniceus Henderson, 1896
| Mclaughlin, Patsy A. 2004 |
Paguristes puniceus
| Miyake 1978: 38 |
Paguristes puniceus
| Balss 1912: 92 |
Paguristes puniceus
| Komai 2001: 423 |
| Gordan 1956: 323 |
| Balss 1924: 768 |
| Kemp 1912: 25 |
| Alcock 1905: 38 |
| Henderson 1896: 527 |