
Tropidophis cacuangoae, Ortega-Andrade & Bentley & Koch & Yánez-Muñoz & Entiauspe-Neto, 2022
|
publication ID |
https://doi.org/ 10.5852/ejt.2022.854.2021 |
|
publication LSID |
lsid:zoobank.org:pub:8EA03380-77C9-4DA8-9A4E-9005D19D91E4 |
|
DOI |
https://doi.org/10.5281/zenodo.7525729 |
|
persistent identifier |
https://treatment.plazi.org/id/03A88798-2745-B530-FDD9-FBC73049FCA5 |
|
treatment provided by |
Felipe |
|
scientific name |
Tropidophis cacuangoae |
| status |
sp. nov. |
Tropidophis cacuangoae sp. nov.
urn:lsid:zoobank.org:act:BF8BCF5A-B93E-4C89-A9C5-EAE47B662863
Figs 1 View Fig , 4–13 View Fig View Fig View Fig View Fig View Fig View Fig View Fig View Fig View Fig View Fig , 16 View Fig
Diagnosis
Tropidophis cacuangoae sp. nov. can be distinguished from all its congeners based on the following combination of characters: (1) slightly keeled dorsal scales with no apical pits, 23/22/17 or 22/23/17 rows; (2) postoculars three; (3) loreal absent, possibly fused with nasal plate; (4) temporal formula 3 + 4 + 3 or 2 + 3 + 3; (5) supralabials 8–10, with 4 th and 5 th in contact with eye orbit; (6) infralabials 10–11, with first two pairs in contact with anterior chinshields; (7) ventrals 162 in single male, 156 in single female; (8) subcaudals single, 33 in single male, 30 in single female, male with cloacal spines; (9) in life, dorsal background coloration light brown, with diffuse dorsolateral black blotches, gray vertebral coloration, dark brown postocular stripe and diffuse dark brown pigmentation on labials; (10) in life, ventral background coloration light orange, with large ocellar black blotches, ranging from 16–20, covering up to four ventrals each; (11) maximum SVL 271 mm in single male, and 255 mm in single female; (12) maximum tail length 49 mm in single male, 40 mm in single female; (13) vertebral scale rows 161–162 on body, 32–35 on tail; (14) nasal contacting rostral, first and second supralabials, preocular, anterior prefrontal, and internasal; (15) two pairs of prefrontals; (16) maxillary teeth 18‒21; (17) prefrontal and maxilla bones in contact; (18) dentary and angular bones in contact; (19) medial contact of exoccipitals loose; (20) small ventral ethmoidal foramen in premaxilla body; (21) cervical vertebrae two or less; (22) vomer further expanded laterally to completely encapsulate vomeronasal organ posteriorly; (23) postorbital, dorsomedial head divided in two heads.
Etymology
The specific epithet ʻ cacuangoae ʼ, is a noun in genitive case, a Latinization honoring Dolores Cacuango, an Ecuadorian benchmark of feminism and human rights of the early twentieth century. She claimed the identity and rights of the Ecuadorian indigenous people, leading them to defend themselves from abuse and discrimination. Also, she demanded the teaching of Quechua and founded the first bilingual schools in Ecuador and the Ecuadorian Indigenous Federation.
Type material
Holotype ECUADOR • adult ♂; Napo Province, San Juan de Muyuna Municipality, Reserva Biológica Colonso Chalupas , Campamento 2 ; 0.93370 N, 77.92239 W; alt. 1613 m; H.M. Ortega-Andrade leg.; sequence: DHMECN 16725 ; DHMECN 16725. ( Figs 4 View Fig , 7A–C View Fig , 8A View Fig , 16 View Fig ). GoogleMaps
Paratype ECUADOR • adult ♀; Pastaza Province, Mera County, Sumak Kawsay In Situ reserve ; 1.3930470 N, 78.0686059 W; alt. 1481 m; A. Bentley leg.; sequence: HMOA1935_RBCC; DHMECN 15893 . ( Figs 5–7, 7D–F View Fig View Fig View Fig , 8B View Fig , 9–13 View Fig View Fig View Fig View Fig View Fig ) GoogleMaps .
Referred specimen
ECUADOR • adult ♂; Zamora-Chinchipe Province, Guayzimi , Nangaritza Municipality; sequence: AB161_NTF3; DHMECN 16126 . Unconfirmed candidate new species ( Tropidophis cf. cacuangoae ) .
Description of holotype
Adult male. Total length 320 mm; SVL 271 mm; tail length 49 mm (15.3% of total length, 18.8% of SVL). Head length 8.71 mm (2.72% of total length, 3.21% of SVL); head width 6.78 mm (77% of head length, 2.12% of total length); interorbital distance 3.7 mm (54.5% of head width); rostro-orbital distance 4 mm; nostril-orbital distance 4 mm (45.92% of head length); cervical constriction distinct, with a conspicuously flat vertebral row; head distinct from neck, triangular in dorsal view, narrow anteriorly, arched in lateral view; pupil elliptical, oval shaped; rostral rounded, 2.07 mm wide, projected over lower jaw, length of portion visible in dorsal view slightly smaller than internasal suture; internasals paired, square, 1.45 mm long, 1.32 mm wide; each internasal contacting nasal, rostral, and anterior prefrontal, separated from supralabials; anterior prefrontals paired, square, 1.18 mm long, 1.78 mm wide; each anterior prefrontal contacts internasals, nasal, preocular, and posterior prefrontals; posterior prefrontals paired, square, 0.8 mm long, 1.44 mm wide; each posterior prefrontal contacts frontal, preocular, anterior prefrontals and supraoculars; frontal hexagonal, 3.13 mm long, 1.73 mm wide, contacting posterior prefrontals, supraoculars, and parietals; supraocular trapezoidal, 2.64 mm long, 1.25 mm wide, in contact with preocular, posterior prefrontal, frontal, parietal, and upper postocular; parietals paired, 3.8 mm long, 3 mm at its largest width, broadly in contact, with a single interparietal, contacting parietals, upper postocular, frontal, temporals, and occipitals; occipitals square-shaped, enlarged, in four rows, contacting dorsals; nasal trapezoidal, undivided, 3 mm long, 0.85 mm high, contacting rostral, internasals, 1 st and 2 nd supralabials, anterior prefrontal, and preocular; nostril located in anterior third of nasal, not visible from above; preocular trapezoidal, 0.97 mm long, 1.61 mm high, contacting nasal, anterior prefrontal, posterior prefrontal, 2 nd and 3 rd supralabials, and supraocular; upper postocular 1.03 mm long, 0.98 mm high, contacting supraocular, parietal, middle postocular, and 1 st row of temporals; middle postocular 0.89 mm long, 0.77 mm high, contacting upper postocular, 1 st row of temporals, and lower postocular; lower postocular 0.9 mm long, 0.5 mm high, contacting 5 th, 6 th, and 7 th supralabials, and middle postocular; temporals 3 + 4 + 3, first row contacting upper and middle postoculars, parietal, 7 th and 8 th supralabials, second row contacting parietals, 8 th and 9 th supralabials, third row contacting parietals, occipitals, 9 th and 10 th supralabials; supralabials ten, 4–5 entering orbit; first supralabial contacts rostral and nasal, second contacts nasal and preocular, third contacts preocular, fifth and sixth contact lower postocular, seventh contacts first row of temporals, middle and lower postoculars, eighth contacts first and second row of temporals, ninth contacts second and third row of temporals, tenth contacts third row of temporals; mental triangular, 0.75 mm long, 2.28 mm wide; anterior chinshields paired, 1.95 mm long, 0.68 mm wide; infralabials ten, first pair separated by mental and anterior chinshields, 1–2 in contact with anterior chinshields, 2–3 in unilateral (right side) contact with posterior chinshields, second and third largest, equal in size; dorsal scales slightly keeled, in 23/22/17 rows; pre-cloacal scale entire, with spines; ventrals 162; single preventral; subcaudals entire, 33; terminal scale acuminate, curved ventrally; vertebral scale rows 162 on body, 35 on tail.
Two cervical vertebrae; trunk vertebrae 158; caudal vertebrae 39; remnants of pelvic bones present ( Fig. 8A View Fig ).
Head background coloration light brown dorsally and laterally, dark brown pigmentation dispersed through center of head, coalescing outwards from suture of internasals, prefrontals, and frontal; rostral, supralabials, nasals and preocular with light brown background coloration, and diffuse dark pigmentation on scale edges; conspicuous postocular dark brown stripe, covering upper and middle postoculars, temporals, and dorsals; dark brown pigmentation on edges of infralabials, and into chinshields and gulars; dorsal background coloration light brown, with ocellar black blotches (mostly confined to edges of dorsals), coalescing outwards from dorsoventral region and ventral blotches; ventral background coloration uniformly cream, with 16 ventral ocelli-shaped black blotches.
Variation
The new species is known solely based on two specimens. Measurements (given in mm) range from: SVL 255–271 (255 in female, 271 in male), tail length 40–49 (40 in female, 49 in male), total length 295–320 (295 in female, 320 in male); head length 8.41–8.71 (8.41 in female, 8.71 in male); head width 5.51–6.78 (5.51 in female, 6.78 in male); rostral length 1.07–1.32 (1.07 in female, 1.32 in male); rostral width 1.53–2.07 (1.53 in female, 2.07 in male); internasal length 1.65–1.83 (1.65 in female, 1.83 in male); nasal length 1.78–3 (1.78 in female, 3 in male); nasal width 0.8–0.85 (0.8 in female, 0.85 in male); anterior prefrontal length 0.92–1.18 (0.92 in female, 1.18 in male); anterior prefrontal width 1.78–2.47 (2.47 in female, 1.78 in male); posterior prefrontal length 0.87–0.9 (0.9 in female, 0.87 in male); posterior prefrontal width 1.13–1.44 (1.13 in female, 1.44 in male); frontal length 3.03–3.13 (3.03 in female, 3.13 in male); frontal width 1.73–1.9 (1.9 in female, 1.73 in male); supraocular length 2.23–2.64 (2.23 in female, 2.64 in male); supraocular width 0.83–1.25 (0.83 in female, 1.25 in male); preocular length 0.74–0.97 (0.74 in female, 0.97 in male); preocular width 1.48–1.61 (1.48 in female, 1.61 in male); upper postocular length 1.03–1.06 (1.06 in female, 1.03 in male); upper postocular width 0.98–1.16 (0.98 in female, 1.16 in male); middle postocular length 0.89–0.91 (0.91 in female, 0.89 in male); middle postocular width 0.77–0.97 (0.97 in female, 0.77 in male); lower postocular length 0.7–0.9 (0.7 in female, 0.9 in male); lower postocular width 0.43–0.54 (0.43 in female, 0.54 in male); mental length 0.66–0.75 (0.66 in female, 0.75 in male); mental width 2.09–2.28 (2.09 in female, 2.28 in male); anterior chinshield length 1.92–1.95 (1.92 in female, 1.95 in male); anterior chinshield width 0.67–0.68 (0.67 in female, 0.68 in male); posterior chinshield length 2.32–2.61 (2.61 in female, 2.32 in male); posterior chinshield width 0.75–0.85 (0.75 in female, 0.85 in male). Scale counts may vary: dorsals 23/22/17 or 22/23/17; temporals 3 + 4 + 3 or 2 + 3 + 3; supralabials 8/8 or 10/9, with 3–5 or 3–4 contacting orbit; infralabials 9/9 or 10/9, with 1–2 contacting anterior chinshields; gular scale rows 7–12; vertebral scales on body 161–162, on tail 32–35; ventrals 156–162 (156 in female, 162 in male); subcaudals single, 30–33 shields (30 in female, 33 in male).
The female paratype has two cervical vertebrae, 155 trunk vertebrae and 30 caudal vertebrae, and lacks pelvic remnants ( Fig. 8B View Fig ).
Coloration
The two known specimens largely agree in coloration patterns. Head background coloration light brown dorsally and laterally, dark brown pigmentation dispersed through the center of the head, coalescing outwards from suture of internasals, prefrontals, and frontal; rostral, supralabials, nasals and preocular with light brown background coloration, and diffuse dark pigmentation on scale edges; conspicuous postocular dark brown stripe, covering upper and middle postoculars, temporals, and dorsals; dark brown pigmentation on edges of infralabials, and into chinshields and gulars; dorsal background coloration light brown or light orange, with ocellar black blotches (mostly confined to the edges of dorsals), coalescing outwards from dorsoventral region and ventral blotches; ventral background coloration uniformly cream in preserved specimens, and uniformly orange in life, with 16–20 ventral ocelli-shaped black blotches.
Cranial morphology ( Figs 9–13 View Fig View Fig View Fig View Fig View Fig )
An overview of interspecific osteological comparisons can be seen in Table 1 View Table 1 . Cranial information is mainly based on CT scan data of the paratype DHMECN 15893, an adult female. The skull shows slight damage. The tooth row of the left palatine is missing, the right ectopterygoid and pterygoid are broken and the right quadrate is dislocated. Some information on intraspecific variation is supplemented from CT scan data of the male holotype (DHMECN 16725). The premaxilla is edentulous, subtriangular in frontal view, 2.5 (DHMECN 15893) to 2.8 (DHMECN 16725) times as broad as high, with a short, dorsally oriented ascending process; ascending process almost contacting anterior end of nasals; transverse processes are laterally oriented, 1.3 (DHMECN 16725) to 1.7 (DHMECN 15893) times as long as ascending process, define widest part of premaxilla, entirely visible in dorsal view, posteriorly contact (DHMECN 15893) or not (DHMECN 16725) anteriormost tips of septomaxillae, laterally approach maxillae but remain distinctly separated from them; in ventral view, the posteriorly oriented vomerine process is bifurcate with slightly posterolaterally diverging arms (DHMECN 16725), or tripartite with parallel arms, laterally longer and wider, with rounded ends, and a short and narrower, thumb-like, median arm (DHMECN 15893), lateral arms of each specimen distinctly overlapping anterior part of vomers; ventral surface of premaxilla pierced by two foramina in anterior region.
Nasals are paired, in contact medially, each with an elongate triangular dorsal lamina, which is 3.4 (DHMECN 15893) to 4 (DHMECN 16725) times as long as wide, with a pointed anterior tip and a broadened caudal end, which is laterally slightly overlapped by medial process of prefrontal; posteroventral process of nasal contacts or not anteroventral part of frontal; vertical lamina of nasal contacts medial edge of septomaxilla; nasals are approximately half as broad as septomaxillae and frontals.
Septomaxillae are paired and slightly separated, 1.6 (DHMECN 16725) to 1.9 (DHMECN 15893) times as long as wide, each with a broad ascending conchal process that extends freely anterolaterally beyond lateral margins of nasals but does not reach level of lateral nasal edge; anterior end of septomaxilla touches (DHMECN 15893) or not (DHMECN 16725) lateral process of premaxilla; posterior end remains distinctly separated from anterior end of frontal; ventrally, septomaxilla is connected to vomer; dorsally, connected to vertical lamina of nasal along its entire length, with caudal end being squared off and fitting into a notch in ventral edge of vertical lamina of nasal.
Vomers are paired, complex structures, about three times as long as wide, medially in contact and with anterior region diverging; body of vomer with globular mesoventral portion with an anteriorly oriented opening; anteriorly broad contact zone with dorsal surface of vomerine process of premaxilla; dorsally connected to septomaxilla in anterior and central region; vertical lamina of caudal end of vomer is unforked and dorsolaterally in firm contact with anterior region of choanal process of palatine.
The braincase accounts for approximately 70% of the total skull length.
Prefrontals form anterior margin of orbits; orbital lamina is slightly concave and divided into two bones (DHMECN 15893) or not divided (DHMECN 16725), with lacrimal duct enclosed (DHMECN 16725) or not being enclosed by prefrontal but framed dorsally, laterally, and medially by the two bones (DHMECN 15893); in lateral view, anterior process is convexly rounded; in contact with nasal dorsomedially, frontal posterodorsally, maxilla ventrally, and palatine ventromedially; in posterior view, prefrontal exhibits a small process directed medially or slightly anteromedially.
Frontals are paired, in close medial contact with a straight suture, about semicircular in dorsal view, 1.9 (DHMECN 15893) to 2 (DHMECN 16725) times as long as wide; forming dorsal and medial margins of orbits, slightly convex in dorsal view; anterior margin slightly (DHMECN 15893) or strongly (DHMECN 16725) convexly curved; anterolateral edge of frontal forms a short, thumb-like process that fits into a facet of prefrontal; both frontals together form a rounded suture with parietal, which seamlessly adapts shape of parietal; ventral edges of orbital laminae of frontals in medial contact along their entire length; anteromedially, frontal has a vertical lamina that is fused ventrally with lateral lamina to form a short tubular structure in anterior region of each frontal, with vertical laminae of both frontals being in close medial contact; frontal anteriorly in contact with nasal or slightly separated from it, anterolaterally in contact with prefrontal, posteriorly with parietal, posterolaterally with postorbital (DHMECN 15893) or remains slightly separated from it (DHMECN 16725), ventrally with parabasisphenoid rostrum, and approaching but not touching choanal process of palatine.
Parietal is single, about 1.2 (DHMECN 16725) to 1.4 (DHMECN 15893) times as wide as long, with concavely rounded anterior and anterolateral edges, and convexly rounded lateral and posterior edges in dorsal view; dorsal surface is slightly convex except for a slight longitudinal depression along its midline; not participating in formation of orbit; anterolateral edges in broad and seamless contact with postorbitals; in dorsal view, moderately defined dorsolateral ridges extending posteromedially from posterior point of contact with postorbitals to suture with supraoccipital, approaching but not touching each other; laterally, parietal slopes down forming a strongly convex lateral surface; parietal contacts parabasisphenoid ventrally with an oblique straight suture, frontals anteriorly, prootics posterolaterally with a slightly concave curved suture in dorsal view, and supraoccipital posteriorly with a convex curved suture; parietal does not participate in formation of maxillary branch foramen of trigeminal nerve.
Postorbitals are well developed, elongate, downwardly curved, and meet (DHMECN 15893) or remain slightly separated (DHMECN 16725) from ectopterygoids, in lateral view with a concave anterior margin, forming posterolateral edge of orbit; postorbital contacts parietal posterodorsally, and contacts (DHMECN 15893) or approaches frontal anteromedially without touching (DHMECN 16725).
Supraoccipital is single, subpentagonal, 1.7 (DHMECN 16725) to 1.9 (DHMECN 15893) times as broad as long, with a slightly concave curved anterior suture with parietal, a broadly V-shaped posterior suture with exoccipitals, and an almost straight suture with prootics laterally; lateral edges of supraoccipital are partially overlapped by supratemporals; two lateral ridges originate at anterior margin of supraoccipital lateral to midline, forming a continuation of ridges of parietal, first running parallel to each other for a short distance and then becoming more pronounced and diverging obliquely until they reach posterolateral corner of supraoccipital; in center of supraoccipital there is a medial ridge of about one third (DHMECN 15893) or half (DHMECN 16725) of length of supraoccipital; internally, lateral edges of supraoccipital extend downward and contribute to dorsomedial walls of otic capsules.
Exoccipitals are paired, irregularly shaped, dorsally in loose medial contact along a straight suture; each has a moderately pronounced dorsal ridge that continues corresponding lateral ridge of supraoccipital; each exoccipital contacts supraoccipital anterodorsally, prootic anterolaterally, supratemporal dorsolaterally, and basioccipital ventrally; large fenestra ovalis is located at suture of prootic and exoccipital, the latter forming posterior margin of fenestra; exoccipitals form dorsal, lateral, and lateroventral border of foramen magnum, without preventing basioccipital from its participation in foramen magnum; internally, exoccipital contributes to formation of posteroventral, posteromedial, and posterolateral walls of the otic capsule.
Columella has an enlarged ovaloid footplate inserted into foramen ovalis and firmly in contact with prootic anteriorly and with exoccipital posteriorly; long and thin, posterolaterally oriented stylus extends toward inner surface of quadrate but remains separate from it.
Prootics are ovaloid in lateral view with an uneven surface, 1.2 (DHMECN 16725) to (DHMECN 15893) 1.4 times as high as long; each has a large anterior and posterior trigeminal foramen separated by laterosphenoid, anterior foramen lying entirely within prootic and not at suture with parietal; each prootic contacts parietal anteriorly and dorsoanteriorly, supraoccipital dorsally, exoccipitals posteriorly, parabasisphenoid complex anteroventrally, basioccipital posteroventrally, and is partially overlapped by supratemporal dorsally; posteriorly, prootic forms anterior margin of fenestra ovalis at suture with exoccipital; internally, prootic contributes to formation of anteroventral, anteromedial and anterolateral walls of otic capsule.
Basioccipital is pentagonal and ends posteriorly in a blunt, rounded tip; about 1.2 times as wide as long, with its widest part just posterior to suture of prootic and exoccipital; ventral surface of basioccipital is convex; it contacts parabasisphenoid complex anteriorly with a slightly undulate suture, prootics anterolaterally with an oblique straight suture, exoccipitals posterolaterally with an oblique straight suture, and forms ventral border of foramen magnum posteriorly; it is anteroventrally overlapped by pterygoids (DHMECN 16725) or remains slightly separated from them (DHMECN 15893); a short slit-like depression is absent (DHMECN 16725) or located centrally on anterior margin of basioccipital (DHMECN 15893).
Parasphenoid and basisphenoid are fused to form an elongate parabasisphenoid that is about twice as long as wide, with its widest part at contact zone of parietal and prootics; parasphenoid rostrum intervenes ventrally between vertical laminae of frontals and is very narrow and long, with a pointed tip that ends anteriorly well in front of anterior margin of frontals; parasphenoid rostrum laterally contacts (right palatine of DHMECN 15893) or not (DHMECN 16725, left palatine of DHMECN 15893) choanal process of palatine, and does not contact nasals, vomers or septomaxillae; parasphenoid rostrum exhibits a lateral groove at its base on each side; parabasisphenoid contacts parietal laterally, prootics posterolaterally and basioccipital posteriorly; on each side of basisphenoid at about midpoint of suture with prootic, there is a short elongated groove directed perpendicular to suture, which continues approximately with same length on prootic.
Maxillae are long, account for 45% (DHMECN 16725) to 46% (DHMECN 15893) of skull length, and are about 8 (DHMECN 15893) to 9.8 (DHMECN 16725) times as long as high, with the highest point located at about the level of the fourth tooth; each extending from level of anterior end of vomer to posterior border of postorbital; posterior half forming major part of lower lateral margin of orbit; anteriorly, slightly curved towards premaxilla; ventral surface of maxilla bears 18‒21 tooth loci; teeth are slightly higher in anterior region, curved, and rear-facing; a moderately long, subrectangular palatine process is located at about mid-region of maxilla at level between 9 th and 12 th tooth, facing posteromedially and slightly curved in ventral direction; a short, knob-like medially oriented ectopterygoid process is located at level of the penultimate and antepenultimate teeth, slightly anterior to anterior tip of medial process of ectopterygoid; each maxilla contacts prefrontal dorsally at two locations both just anterior to and at level of palatine process, maxillary process of palatine dorsomedially by means of palatine process, and lateral process of ectopterygoid posteriorly.
Ectopterygoids are elongate, 5.5 (DHMECN 16725) to 5.6 (DHMECN 15893) times as long as wide, with widest point at level of medial process, approximately axe-shaped in dorsal view; lateral and medial processes are almost indistinguishable, posterior process is long, very narrow, and rod-shaped; in dorsal view, only anterior region of ectopterygoid is visible; each ectopterygoid contacts dorsoposterior end of maxilla with its lateral process, dorsal surface of middle region of pterygoid with posterior half (DHMECN 15893) or one-third (DHMECN 16725) of its posterior process, and it touches (DHMECN 15893) or not (DHMECN 16725) ventral end of postorbital with its lateral process at level of contact point with maxilla.
Palatines are robust, almost straight in ventral view, and account for just under one-third of skull length, each has a moderately long, posterolaterally oriented maxillary process, located about mid-length at level of 5 th (DHMECN 15893) or 6 th (DHMECN 16725) tooth, and a very large and broad choanal process, extending from about level of maxillary process to posterior end of palatine; choanal processes of both palatines approach each other to a very short distance; ventral surface with 7‒9 tooth loci; teeth about similar in size, curved, and rear-facing; maxillary process contacts (DHMECN 15893) or not (DHMECN 16725) palatine process of maxilla ventrally and touches prefrontal dorsally; choanal process contacts posterior region of vomer medially at its anterior region, and touches (right palatine of DHMECN 15893) or not (DHMECN 16725, left palatine of DHMECN 15893) parasphenoid rostrum dorsally; posteriorly, palatine contacts anterior region of pterygoid; in dorsal view, maxillary processes are the only visible part of palatines.
Pterygoids are elongate, and account for about 60% of skull length; ventral surface bears 11–13 tooth loci; teeth subequal, slightly decreasing in size posteriorly, curved, and rear-facing; each pterygoid is narrowed in anterior part up to level of 8 th to 11 th tooth and then broadens abruptly at level of contact zone with ectopterygoid, about last quarter tapers sharply and is posterolaterally oriented; anterior region is dorsally overlain by posterior portion of palatine; pterygoids are slightly (DHMECN 16725) or moderately (DHMECN 15893) curved along tooth line, and have the greatest distance from each other at their posterior ends; posterior end of pterygoid approaches medially ventromedial process of quadrate without touching it; apart from palatine and ectopterygoid, pterygoid touches basioccipital (DHMECN 16725) or remains separated from it (DHMECN 15893); dorsal surface behind tooth line is concavely curved; in dorsal view, pterygoids are hardly visible, being mainly covered by roofing skull bones.
Supratemporals are laminar, elongate, and broad, about 2.3 (DHMECN 15893) to 2.6 (DHMECN 16725) times as long as wide, and exhibit their greatest width at about level of contact zone of prootic and exoccipital; anterior half of each supratemporal overlaps and firmly attaches to prootic, ending well before suture of prootic with parietal; anterior portion of posterior half overlies anterolateral portion of exoccipital, while most of posterior half extends freely and surpasses exoccipital and quadrate posteriorly, forming caudal margin of skull; supratemporal overlaps supraoccipital medially and approaches quadrate with its posterolateral surface, but remains slightly separated from it.
Quadrates are flattened and broad dorsally, about 1.4 times as high as long, oriented vertically and obliquely from anterodorsal to posteroventral, and tapering dorsoventrally in lateral view; no short process is visible in medial part; ventral part is slightly bifurcated and spans glenoid cavity of retroarticular process of compound bone; quadrate does not exceed posterior limit of skull roof.
Each mandible consists of dentary, splenial, angular, compound bone, and a tiny coronoid; in dorsal view, it is almost completely straight, except for anterior third of dentary, which is curved medially ( Fig. 13 View Fig ).
Dentaries are elongate, account for about 55% of lengths of mandibula; dorsal surface of each dentary bears 22‒25 tooth loci; teeth are subequal, curved, rear-facing, and slightly decreasing in size posteriorly; elongate mental foramen is located at about level of 9 th or 10 th tooth; at about level of 13 th (DHMECN 15893) or 15 th (DHMECN 16725) tooth, dentary branches into a long, slender, toothed dorsal process that overlies compound bone and contacts or not coronoid medially, and a ventromedial process about half as long or even shorter that is in contact with splenial for almost its entire length, and loosely connected to anterior portion of compound bone; at level of 16 th (DHMECN 15893) or 19 th (DHMECN 16725) tooth, dorsal process branches again into a tiny knob-like medial process and a much longer tooth-bearing dorsal process; inner surface bears a long groove in extension to depression for splenial, which extends anteriorly to level of 2 nd tooth.
Coronoids are tiny, elongate, about 4.5 times as long as high, and by far the smallest bones of the mandibles; each contacts dentary anteriorly and dorsolaterally or remains slightly separated from dentary, compound bone laterally, and angular ventrally (DHMECN 16725) or remains slightly separated from it (DHMECN 15893).
Splenials are elongate, triangular, tapering anteriorly to a sharp tip, 1.6 (DHMECN 16725; due to posterior dorsal extension) or 5.5 (DHMECN 15893) times as long as high, with (DHMECN 16725) or without (DHMECN 15893) a dorsal extension at posterior end; anterior mylohyoid foramen is present in posterior region; each extends medially along dentary between 10 th and 17 th tooth (DHMECN 15893) or between 13 th and 20 th tooth (DHMECN 16725), and posteriorly contacts anterior region of angular, and has a minimal contact point with compound bone.
Angulars are elongate, triangular, tapering posteriorly to a blunt (DHMECN 15893) or pointed (DHMECN 16725) tip, and about twice (DHMECN 16725) or 3 times (DHMECN 15893) as long as high, accounting for 77% (DHMECN 15893) to 80% (DHMECN 16725) of length of splenial; each angular contacts compound bone laterally and dorsally, splenial anteriorly, and touches (DHMECN 16725) or not (DHMECN 15893) coronoid anterodorsally; a posterior mylohyoid foramen is absent (DHMECN 15893) or slightly indicated (DHMECN 16725).
Compound bones are elongate, about 6.4 times as long as high, and account for about two-thirds of length of mandible; each attaches to dentary at level of 14 th (DHMECN 15893) or 16 th (DHMECN 16725) tooth, and in lateral view its anteriorly tapering rostral end extends between dorsal and ventromedial processes of dentary; medial prearticular crest is prominent, completely visible in lateral view, much higher than surangular crest, the latter not visible in medial view; anteriorly oriented foramen is located at or just behind point where posterior tip of toothed dorsal process of dentary meets compound bone; mandibular fossa is deep, elongate, narrow, dorsolaterally oriented and visible in lateral view; articular part, where it is joined to quadrate, has a deep, saddle-shaped notch; retroarticular process is short, slightly directed medially, and does not extend beyond posterior end of exoccipital in dorsal view.
Comparisons
Characteristics from other species are presented in parentheses in this section. Comparisons are restricted to mainland tropidophiid species (sensu Curcio et al. 2012). An overview of morphological comparisons can be seen in Table 2 View Table 2 . In its geographic range, Tropidophis cacuangoae sp. nov. might only be confused with another andine amerophid, Tropidophis taczanowskyi (Steindachner, 1880) ( Figs 14–16 View Fig View Fig View Fig ), which occurs in Ecuador and Peru. Both species share a small-sized and stout body, and similar counts of ventrals, subcaudals, supralabials, and infralabials. However, there are striking differences between these species. The dorsal scales of Tro. cacuangoae sp. nov. have lower counts for the anterior and posterior rows, ranging from 22/23/17 to 23/22/17 (23/23/21, 23/23/19, 25/23/ 21 in Tro. taczanowskyi ), and are also mostly smooth, with small or indistinct keels (conspicuously keeled in Tro. taczanowskyi ). The new species also has different temporal scale formulae, ranging from 2 + 3 + 3 to 3 + 4 + 3 (2 + 3 in Tro. taczanowskyi ), and absent or small interparietals (present in Tro. taczanowskyi ), vertebral row enlarged (not enlarged in Tro. taczanowskyi ), retaining contact between both parietals (usually absent in Tro. taczanowskyi ). Furthermore, these two species also have significant differences in coloration and pattern, as Tro. cacuangoae sp. nov. has a dorsal pattern uniformly light brown, with small diffuse brown spots (striped-spotted, lateral orange stripes over a dark brown background in Tro. taczanowskyi ), a prefrontal and maxilla bone contact present (absent in Tro. taczanowskyi ), a dentary and angular bone contact present (absent in Tro. taczanowskyi ), medial contact of exoccipitals loose (firm in Tro. taczanowskyi ), a small premaxilla body ventral ethmoidal foramen (large in Tro. taczanowskyi ), postorbital dorsomedial head divided in two heads (undivided in Tro. taczanowskyi ), vomer further expanded laterally to completely encapsulate vomeronasal organ posteriorly (with margins enclosing posterior wall sloping ventrolaterally in Tro. taczanowskyi ), fenestra ovalis oriented posterolaterally (fenestra ovalis opens ventrolaterally in Tro. taczanowskyi ), frontal descending process abuts parietal (parietal overlaps frontal laterally in Tro. taczanowskyi ), and ventral pattern with large black blotches on an orange background (alternating black squares on a white background in Tro. taczanowskyi ).
Another tropidophiid species reported from the Andes range is Tropidophis battersbyi Laurent, 1949 . This species was described based on a specimen received by The Royal Museum of Natural History in Brussels in 1874–1875, from an unknown locality in Ecuador. The species is known solely based on its holotype, and no other available information has surfaced. Díaz & Cádiz (2020) highlight that Tro. battersbyi likely represents a labeling error, and could not be diagnosed from two Antillean taxa, Tropidophis spiritu s Hedges & Garrido, 1999 and Tropidophis morenoi Hedges, Garrido & Diaz, 2001 . Nonetheless, Tro. cacuangoae sp. nov. can be readily distinguished from Tro. battersbyi based on its uniformly light brown coloration pattern (large dark brown spots over a light gray background in Tro. battersbyi ), its lower ventral scale counts, which range from 156 to 162 (200 in Tro. battersbyi ), and lower subcaudal counts, which range from 30 to 33 (41 in Tro. battersbyi ).
The genus Trachyboa contains two species, Trachyboa gularis Peters, 1860 and Trachyboa boulengeri Peracca, 1910 , that also occur in Ecuador, and might be confused with the new species. These are also stout bodied, brown colored, small-sized snakes. However, Tropidophis cacuangoae sp. nov. is readily distinguished from these species in having mostly smooth or weakly keeled dorsal scales (rugous and keeled in Tra. gularis and Tra. boulengeri ), a large and entire rostral plate (absent or fragmented into small scales in Tra. gularis and Tra. boulengeri ), a single preocular (fragmented in a circular row of up to 14 ocular scales in Tra. gularis and Tra. boulengeri ), and for lacking horn-like supraocular projections (present in Tra. boulengeri ). However, both species of Trachyboa occur west of the Andes, while Tropidophis cacuangoae occurs east of the Andes.
Within mainland South America, there are three other extralimital species of Tropidophis : Tropidophis paucisquamis (Müller in Schenkel, 1901) ( Fig. 17 View Fig ), Tropidophis grapiuna Curcio, Nunes, Argôlo, Skuk & Rodrigues, 2012 , and Tropidophis preciosus Curcio, Nunes, Argôlo, Skuk & Rodrigues, 2012 . These three species are largely allopatric between each other, and restricted to the Atlantic Forest highlands in southern, southeastern, and northeastern Brazil. The dorsal coloration of Tro. cacuangoae sp. nov. resembles that of Tro. paucisquamis , differing in having a uniformly colored dorsum (small dark brown spots over a dark brown background, vertebral light brown stripe in Tro. paucisquamis ), and this species is furthermore distinguished by its lower ventral counts, which range from 156 to 162 (164–178 in males, 167–183 in females of Tro. paucisquamis ). The new species can be readily distinguished from Tro. grapiuna and Tro. preciosus based on its distinct, uniformly colored dorsal pattern (dark brown with black spots in both species), lower ventral counts, which range from 156 to 162 (196–203 in females of Tro. preciosus ), and lower infralabial counts, which range from 8 to 10 (11–12 in Tro. preciosus and Tro. grapiuna ).
Geographic distribution and natural history
Currently, two verified specimens of T. cacuangoae sp. nov. are known from Ecuador, one (DHMECN 16725) from Reserva Biológica Colonso Chalupas, Campamento 2 (0.93370° N, 77.92239° W), San Juan de Muyuna Municipality, 1613 m altitude, Napo Province, and the other (DHMECN 15893) from Sumak Kawsay In Situ reserve (1.3930470° N, 78.0686059° W), Mera County, 1481 m altitude, Pastaza Province ( Fig. 18 View Fig ). Another specimen is similar to T. cacuangoae sp. nov. (DHMECN 16126), from Guayzimi, Nangaritza Municipality, in Zamora-Chinchipe Province, Ecuador, and recognized herein as ʻUnconfirmed Candidate Species 1ʼ ( Tropidophis cf. cacuangoae ). The new species inhabits primary Eastern Piedmont Evergreen and Evergreen Low Montane forests, under the Amazon Rainforest biome, in Napo and Pastaza provinces, at an altitude range of 1481–1613 m above sea level.
The holotype was collected active during the night (23:36 hours), found in the leaf litter, in Evergreen Low Montane Forest. The paratype was collected during the morning (10:00 hours), found coiled between two wooden boards, on the edge of primary Eastern Piedmont Evergreen Forest. Upon being handled, the specimen did not attempt to bite, and engaged in ʻballingʼ behavior, cloacal discharges and hiding head between body coils. After being handled, the paratype specimen also engaged in autohemorraging of the eyes.





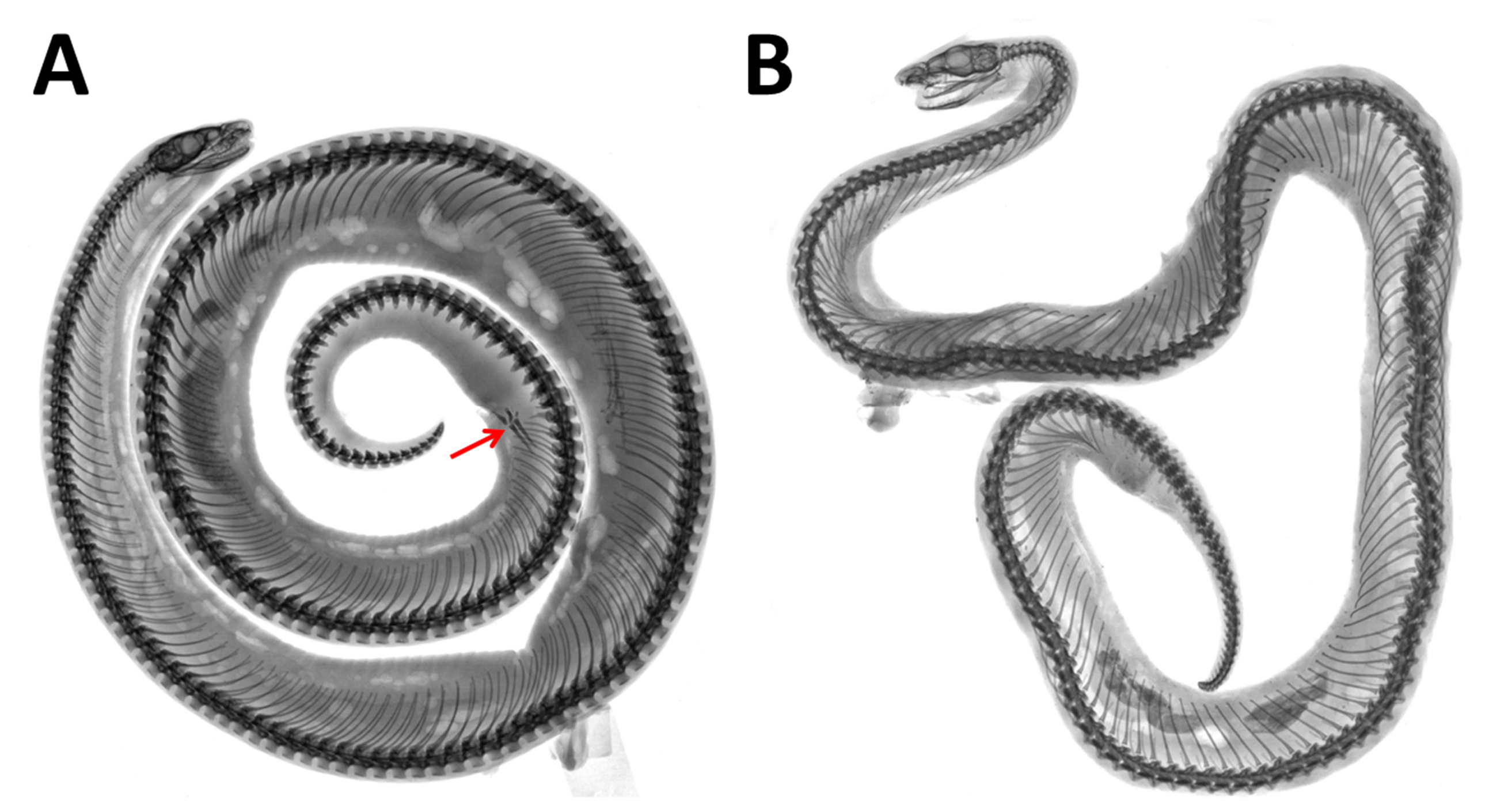
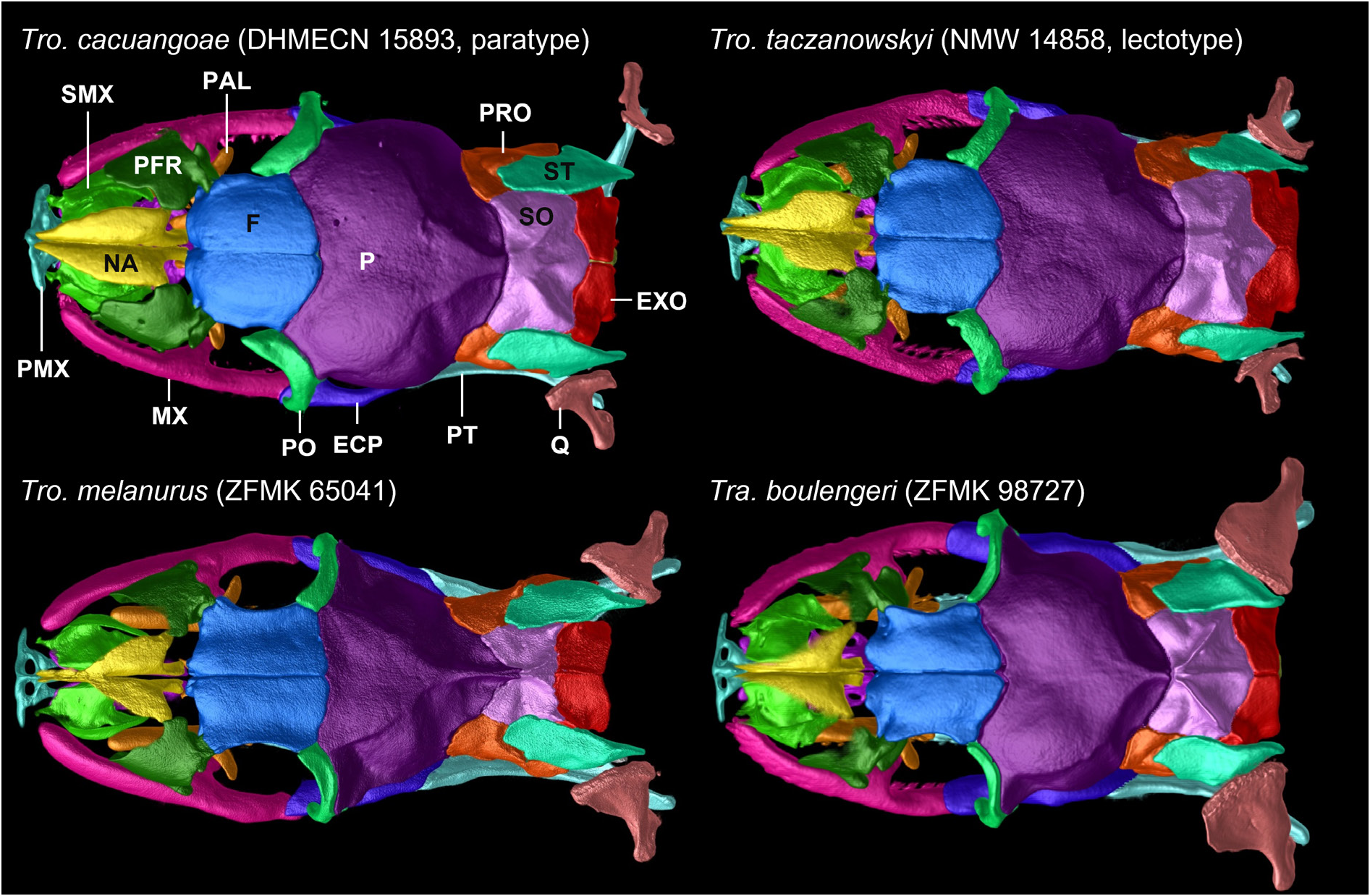


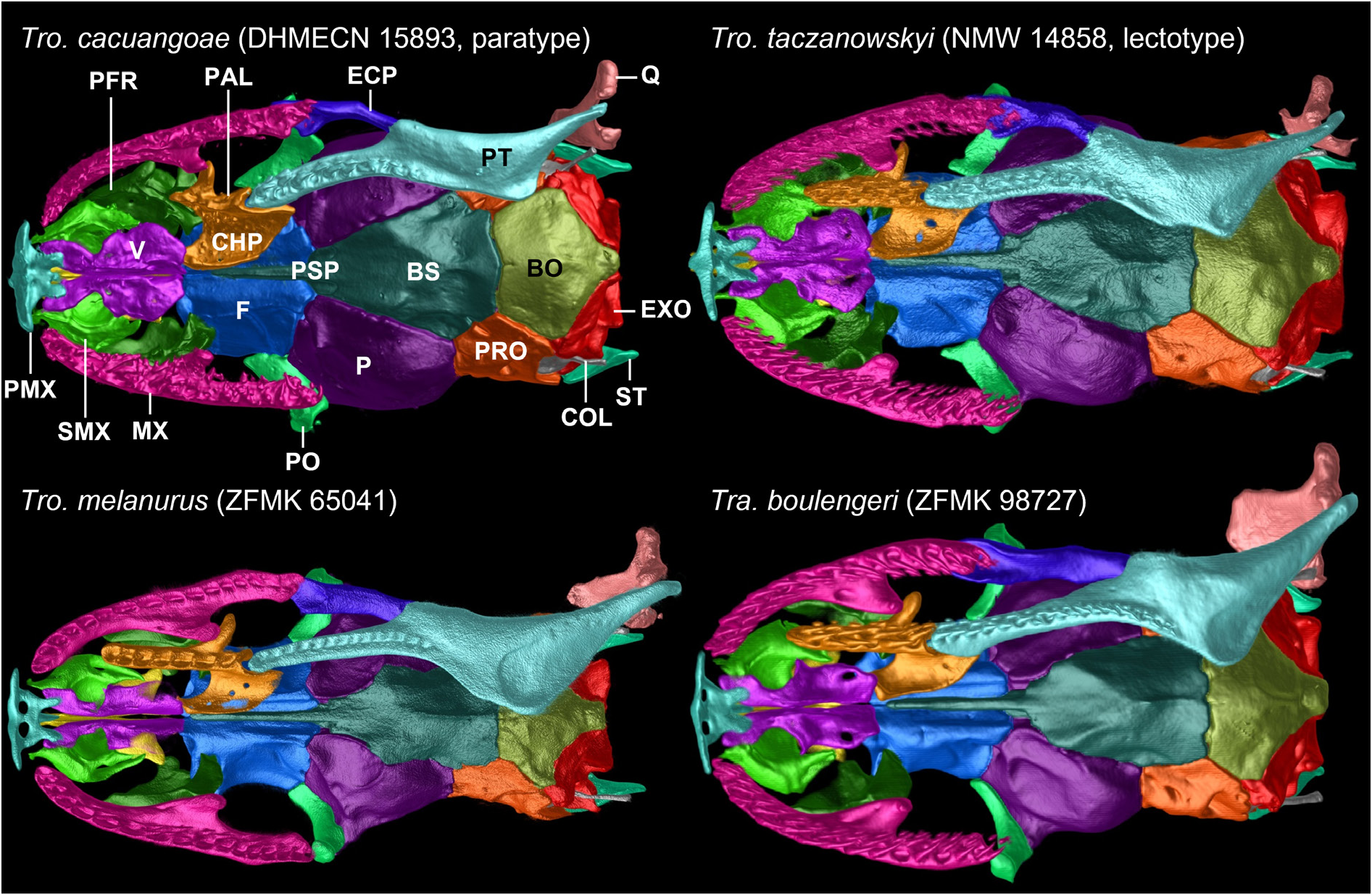
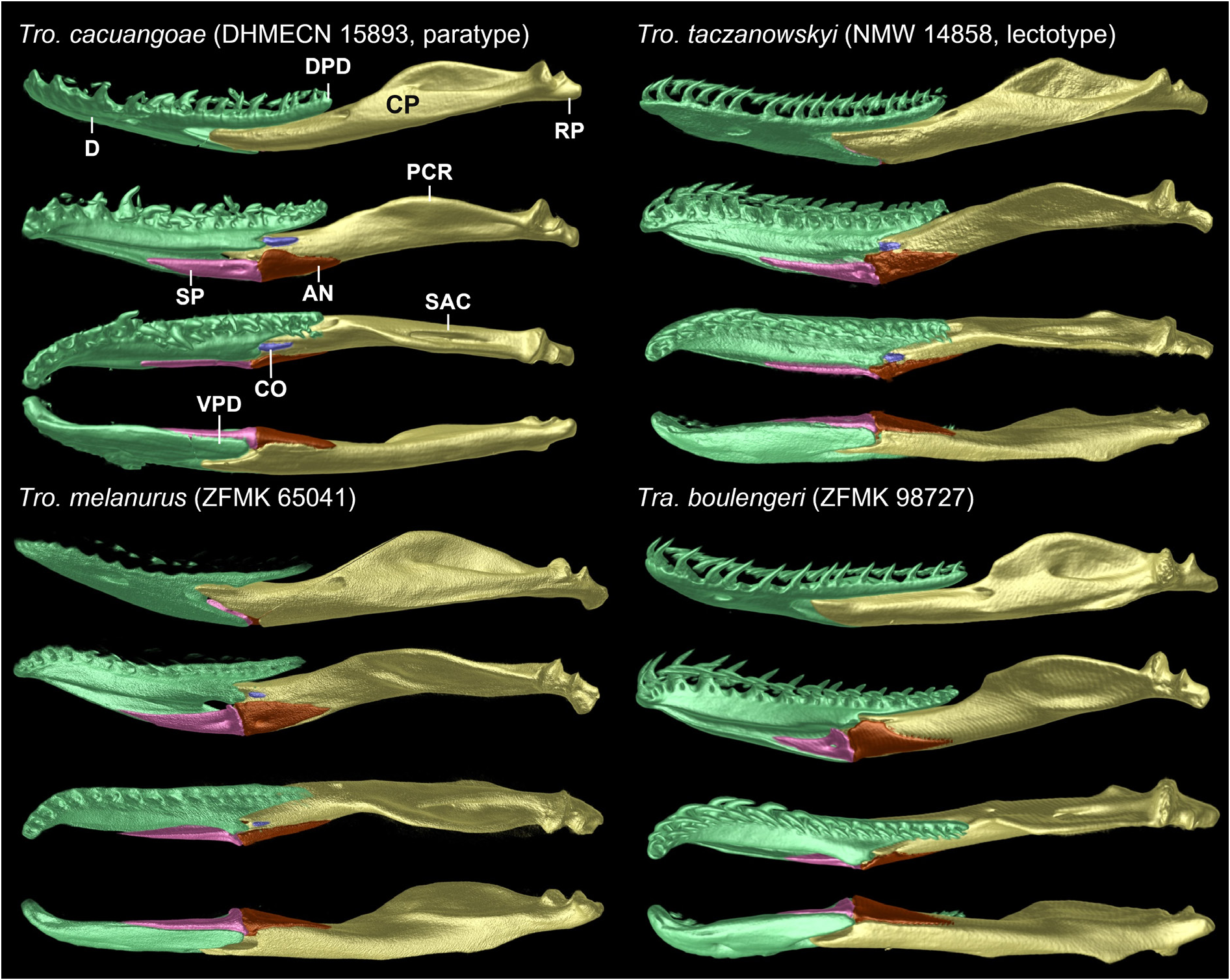
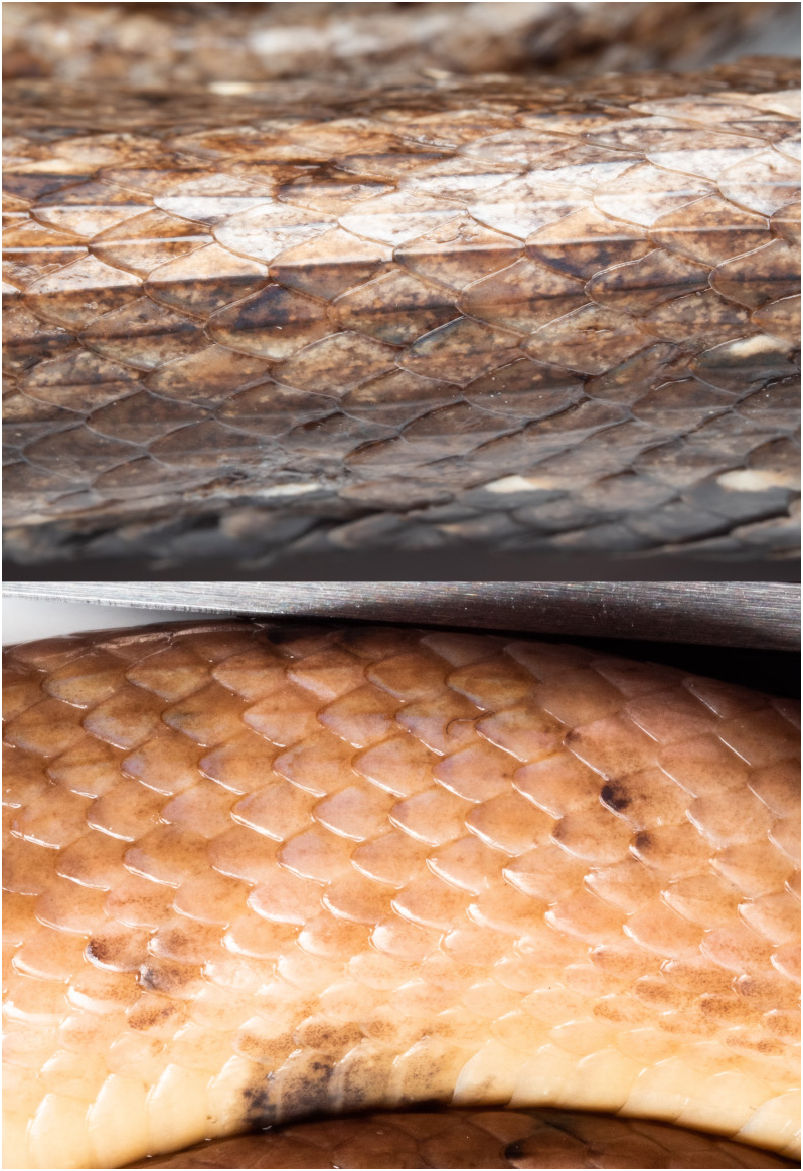




Table 2. Morphological comparisons among mainland Tropidophis Bibron, 1840 of South America. Diagnostic characters of Tropidophis cacuangoae sp. nov. are highlighted in bold. Supporting data from Curcio et al. 2012 are indicated with an asterisk (*). Species with confirmed records from the Andes mountain range are indicated with a dagger (†).
| Tro. cacuangoae sp. nov. † | Tro. battersbyi | Tro. taczanowksyi * † | Tro. grapiuna * | Tro. paucisquamis * | Tro. preciosus * | |
|---|---|---|---|---|---|---|
| Dorsals | 23/22/17 (n = 1) or | 21/23/17 (n = 1) | 23/23/21 (n = 7), | 23/23/19 (n = 1) | 16/21/15 (n = 1), 21/21/15 | 23/23/17 (n = 1) |
| 22/23/17 (n = 1) | 25/23/21 (n = 4), or | (n = 12), 21/21/16 (n = 3), | ||||
| 23/23/19 (n = 2) | 21/21/17 (n = 2), 21/23/17 | |||||
| (n = 2), 23/23/16 (n = | ||||||
| 2), 23/23/17 (n = 14), | ||||||
| 23/23/18 (n = 1), 23/23/19 | ||||||
| (n = 3) | ||||||
| Keels | present, small or indistinct | absent | present, conspicuous | present | absent or indistinct | absent |
| Ventrals | 162 (♂) (n = 1); | 200 (♂) (n = 1) | 148–160 (♂) (n = 4); | 154–155 (♀) (n = 2) | 164–178 (♂) (n = 20); | 196–203 (♀) (n = 2) |
| 156 (♀) (n = 1) | 146–156 (♀) (n = 9) | 167–183 (♀) (n = 30) | ||||
| Rostral scale | present, entire | present, entire | present, entire | present, entire | present, entire | present, entire |
| Temporals | 3 + 4 + 3 (n = 1) or | 3 + 4 (n = 1) | 2 + 3 (n = 13) | 2 + 3 (n = 2) | 2 + 3 (n = 8) | 2 + 3 (n = 2) |
| 2 + 3 + 3 (n = 1) | ||||||
| Subcaudals | 33 (♂) (n = 1); | 41 (♂) (n = 1) | 25–33 (♂) (n = 9); | 26–30 (♀) (n = 2) | 29–38 (♂) (n = 19); 30–37 | 27 (♀) (n = 1) |
| 30 (♀) (n = 1) | 23–27 (♀) (n = 4) | (♀) (n = 29) | ||||
| Supralabials | 8–10 (n = 2) | 11 (n = 1) | 8–11 (n = 13) | 10–11 (n = 2) | 8–10 (n = 12) | 9–10 (n = 1) |
| Infralabials | 9–10 (n = 2) | 11 (n = 1) | 9–11 (n = 13) | 11–12 (n = 2) | 8–11 (n = 12) | 11–12 (n = 2) |
| Dorsal pattern | uniformly light brown, | large dark brown | striped-spotted, lateral | small dark brown | small dark brown spots | irregular dark brown |
| small diffuse brown spots | spots over a light gray | orange stripes over a | spots over a dark | over a dark brown | blotches over a dark | |
| background | dark brown background | brown background | background, vertebral | brown background | ||
| light brown stripe | ||||||
| Ventral pattern | large black blotches on an | alternating black | alternating black | alternating black | large black elliptical | alternating large black |
| orange background | elliptical blotches | squares on a white | elliptical blotches | blotches on an orange | squares | |
| background | background | |||||
| Eyelashes | absent | absent | absent | absent | absent | absent |
| Parietal contact | in contact | separated | usually separated | separated | in contact | in contact |
| Interparietals | absent or small | present, conspicuous | present, conspicuous | present, conspicuous | usually absent | absent |
| Maximum SVL (mm) | 320 (♂), 255 (♀) | 299 (♂) | 335 (♂), 348 (♀) | 289 (♀) | 336 (♂), 357 (♀) | 449 (♀) |
| Geographic | Northeastern Ecuador | Ecuador, but likely | Southeastern Ecuador | Northeastern Brazil | Southeastern Brazil | Southeastern Brazil |
| distribution | from Greater Antilles | and Northeastern Peru. |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
SubOrder |
Serpentes |
|
SuperFamily |
Amerophidia |
|
Family |
|
|
Genus |