
Craniella quirimure, Peixinho, Solange, Cosme, Bruno & Hajdu, Eduardo, 2005
|
publication ID |
https://doi.org/ 10.5281/zenodo.169740 |
|
DOI |
https://doi.org/10.5281/zenodo.6268966 |
|
persistent identifier |
https://treatment.plazi.org/id/03A79171-FFBF-FFA5-BA31-FA0FAC6BFDBF |
|
treatment provided by |
Plazi |
|
scientific name |
Craniella quirimure |
| status |
sp. nov. |
Craniella quirimure sp. nov.
Figs. 2 View FIGURE 2 , 3, 4 View FIGURE 4
Holotype — UFBA 1812POR, (1) Barra do Jacuruna (ca. 13o02.857’S 38o48.857’W, Itaparica Channel, Jaguaripe, State of Bahia, Brazil) intertidal, mangrove, coll. S. Peixinho, i.1976.
Paratypes (1) Barra do Jacuruna (ca. 13o02.857’S 38o48.857’W, Itaparica Channel, Jaguaripe, State of Bahia, Brazil) intertidal, mangrove — UFBA 212POR 15 specimens), coll. A. Sampaio, 02.xi.1975; UFBA 222POR (7 specimens), coll. S.Peixinho, 24.viii.1975; UFBA 252POR (9 specimens), coll. A. Sampaio, iv.1976; UFBA 254POR (3 specimens), coll. S. Peixinho, i.1976; UFBA 262POR, UFBA 161 PGR, Coll. S.
Peixinho, 1976 (49 specimens), UFBA 1811POR, coll. S. Peixinho, 1976; MNRJ 601 (1 specimen), coll. E. Hajdu, S. Peixinho & R. DesqueyrouxFaúndez, 22.i.1997; UFBA 1776POR (8 specimens), coll. S. Peixinho, E. Hajdu, R.G.S. Berlinck, C.P. Santos & E.L. Esteves, 05.vi.2004; MNRJ 8417 (5 specimens), coll. S. Peixinho, E. Hajdu, R.G.S. Berlinck, C.P. Santos & E.L. Esteves, 05.vi.2004. (2) Itaparica Marina (S12º53,340' / W38º41,075', Itaparica Island, State of Bahia, Brazil) UFBA 1888POR (2 specimens), coll. B. Cosme, 30.i.2005. (3) Maraú River Delta (S13º58´/ W38º59´, Bay of Camamu, State of Bahia, Brazil), mangrove UFBA 999POR (3 specimens), coll. M.C. Guerrazzi et coll., leg. U.S. Pinheiro, 13.vi.2004.
Diagnosis
Craniella quirimure sp.nov. is the only Craniella in the Tropical Atlantic with cortical oxeas up to 1000 µm long and 28 µm thick, two categories of protriaenes, anatriaenes which can be larger than 9000 µm and sigmaspires smaller than 15 µm.
Specimens Oxeas Protriaenes Anatriaenes Sigmaspires
Holotype I. main choanosomal I. large rabdome rabdome 6,28,8 12,8 UFBA 0 1812 16712257 4551/9,921,128 14342148 3156/4,25,16,9>9600/4,16,28,6 POR II. cortical large cladi cladome
574839,71166/14,518,728 21,440,448,8 1825,631
III. anisoxeas large cladome 1318,326
407675,3960/3,66,311,1 II. small rabdome
IV. rhizoidal 210492864/11,31,6
419569727/711,817 small cladi
25,346,464,7
small cladome 820,644
Paratype I. main choanosomal I. large rabdome Rabdome 7.79.811.6 UFBA 0 0 262 756.118772343.3/9.920.828.2 1497.51694.61882.8/4.25.97.7>9000/4.45.97.6 POR II. cortical large cladi cladome
597.9801.81041/9.918.433.3 29.636.442 21.728.233.2
III. anisoxeas large cladome 12.919.123.8
463.4603.1758.7/4.96.515.5 II. small rabdome
IV. rhizoidal 368.8494.7798.1/0.91.42.3
441.4699.71124.8/9.715.428 small cladi
14.638.350.1
large cladome 6.222.633.8
Paratype I. main choanosomal I. large rabdome Rabdome 5.28.612.1 UFBA 0 0 254 1598.32067.72704.9/11.118.825.5 1433.72147.73156.5/4.25.16.9>7000/5.96.77.7 POR II. cortical large cladi cladome
521.1842.51220.6/14.220.731.6 21.440.353.5 24.629.935.7
III. anisoxeas large cladome 21.127.433.4
380.5 594.2808.3/3.95.38.5 II. small rabdome
IV. rhizoidal 209.8491.6864.2/11.31.6
520.4718.2881/13.817.425.3 small cladi
25.346.464.7
large cladome 4.97.311
Paratype I. main choanosomal I. large rabdome Rabdome 5.88.412.3 UFBA 0 1811 1362.81718.52489.8/7.416.321.2 1486.51713.91920.9/5.16.37.8>6000/4.35.96.8 POR II. cortical large cladi cladome
493.4687.31019.2/11.816.124.7 3237.741 24.931.341
III. anisoxeas large cladome 13.818.923.5
462.8540.4726.3/3.96.610.7 II. small rabdome
IV. rhizoidal 212.4362.4514.9/1.11.72.1
269466.2637.2/2.911.216.2 small cladi
33136.742.8
large cladome 10.315.122.5
Description
External form irregularly ovoid, elliptical, barrelshaped or nearly spherical. Specimens reach a maximum of 23mm in diameter and 25mm in height. Surface can be flat (more often) or slightly ridged, with or without perforations (inhalant apertures). Some specimens were seen to bear a crown of conulosevolcaniform projections on the rim of their apical side. Texture normally smooth. Consistency varies from easily compressible to nearly incompressible, what might be related to contraction after collection and handling. More often a single apical oscule occurs, also markedly contracted after collection. A few specimens possessed two or three oscula clustered. Aquiferous channels are clearly seen converging to the oscula in live specimens. In many preserved specimens oscula are not visible. The basal portion of the specimens bears rhizoidal extensions for anchoring in the soft sediment. These are variably intertwined, and can reach 73 mm (UFBA1812POR) in length. In some specimens (ovoid and barrelshaped) these extensions clearly project from several locations spread around the base of specimens. In others (elliptical), they appear to be more concentrated. In live specimens these extensions were either buried in the sediment, or entangled under the specimen, penetrating over 12 cm in the sediment. Live color is muddygreenish and muddyyellowish externally, turning brown in ethanol. Sectioned specimens are yellow inside, clearer on the central portion due to increased silica content, and have two conspicuous outer layers of distinct color. The outermost is sediment colored, reddishbrown, and is adjacent to a bright green subsurface layer.
Skeleton
Nearly perfect radial arrangement of conspicuous spicule tracts of larger choanossomal oxeas spiraling from the base towards the surface of the sponge (ca. 3 tracts in 1 cm of surface area), terminating within a palisadelike layer (ca. 1080 mm thick) of smaller cortical oxeas adjacent to, and slightly piercing (ca. 40 µm) the surface. The radial arrangement is somewhat obscured by a dense crisscrossed reticulation of isolated spicules. Aquiferous channels are seen here and there in the choanosome (up to 280 µm across). Protriaenes are piercing the surface abundantly as commonly seen on other tetillids. Sigmaspires are distributed all over the sponge. The basal rhizoidal extensions have abundant intertwined anchoring anatriaenes.
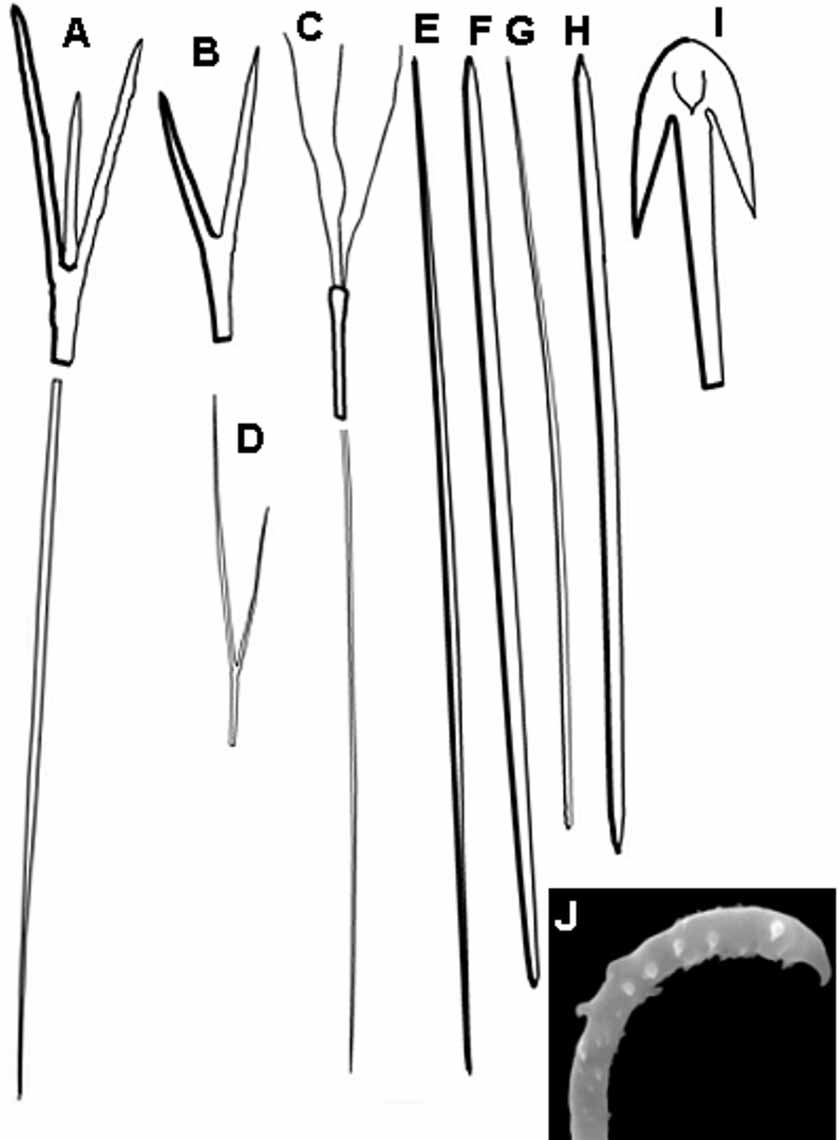
Spicules (measures in m): Megascleres Protriaenes I, associated to the main cortical/ choanossomal tracts, but easier to spot in the cortical region; eventually of prodiaene form; slender. Rhabdome, long, straight, sometimes with a sinuous base, always tapering gradualy; 14341925.93156/4.25.6 7.8 m. Cladome, straight and sharp, ocasionally asymetrical; 2138.754/1320.8 33 m.
Protrianes II, associated to the main cortical/choanossomal tracts, but easier to spot in the cortical region; eventually of prodiaene form; slender. Rhabdome, long, straight, sometimes with a sinuous base, always tapering gradualy; 210473.1908/0.81.4 2.2 m. Cladome, slender, straight or curved, and sharp, ocasionally asymetrical; 1541.965/4.9 16.4 44 m.
Anatriaenes, rhizoidal, forming long tracts embedded in the soft sediment, the cladomes acting as hooks. Extremely long rhabdome, tapering gradually to a sharp end (like a long hair);>6000/46.1 8.5 m. Cladome, straight and sharp 17.628.7 41 m.
Oxeas I, main spicule of the choanosomal bundles; long, straight, stout in the central region and sharpening abruptly, 13631928.12705/717.5 26 m.
Oxeas II, cortical; short and robust, straight, sometimes curved, abruptly sharpened, ocasionally styloid, 493764.81221/1218.3 32 m.
Oxeas III, auxiliary spicules of the choanosomal bundles; anisoxeas, one end tapering gradually, the other abruptly, 381567.2808/3.95.9 11 m.
Oxeas IV, rhizoidal; similar to oxeas I, but shorter, 269592.2881/5.914.3 25 m.
Microscleres Sigmaspires, occurring both in the choanosome as well as in the rhizoidal extensions; small and acanthose, 5.79.0 12 m.
Ecology and distribution
The species is abundant at least on part of the intertidal mangrove area studied (Barra do Jacuruna), where it is found on muddysand substrate attached by its rhizoidal projections. Observed density was about 12 individuals per square meter. Clusters had at most 23 individuals. Water temperature may reach above 30ºC in summer months (DecemberFebruary), and emmersion is calculated to be of no more than 12h on the lowest tydes. This is a delta area where salinity has been reported to vary from 3 to 31/ 1000, with an annual mean around 25/1000 ( Paredes et al., 1980). Other sponges occurring in the area are Mycale microsigmatosa Arndt, 1927 and Haliclona coerulea (Hechtel, 1965) . The Itaparica Marina is situated in a marine environment.
The Bay of Camamu is the target of an ongoing basic inventory of its benthic fauna (M.C. Guerrazzi et coll.). The species was seen on different locations in the bay, at varying distances from the entrance ( U.S. Pinheiro, pers. comm.), which suggests considerable adaptation to salinity fluctuations (not recorded).
Etymology
The species name, a name in apposition, is the Tupinambás Indians name for Todos os Santos Bay, its type locality, and where most of the specimens collected stem from.
Remarks
The very disparate sampling effort undertaken on the three collecting localities precludes any discussion on possible morphologic trends established on these populations. New collections on the Bay of Camamu and on Itaparica Island will allow a study of variability, which is beyond the scope of this contribution.
There are nearly 40 species of Craniella reported worldwide, seven of which from the Tropical Atlantic, viz.: C. carteri Sollas, 1886 ; C. cranium ( Müller, 1789) ; C. insidiosa Schmidt, 1870 ; C. laminaris ( George & Wilson, 1919) ; C. lens Schmidt, 1870 ; C. schmidti Sollas, 1888 and C. tethyoides Schmidt, 1870 .
The new species described here differs from C. carteri and C. lens because both latter species do not possess sigmaspires. Additionally, none of these were reported to possess two categories of protriaenes, and the main choanosomal oxeas of both seem to be always much stouter (always over 20ìm thick) than those of the new species (sometimes as small as 7 m thick). Of those Tropical Western Atlantic Craniella species with sigmaspires, C. schmidti and C. tethyoides have these microscleres much larger (respectively ca. 20 and 35 m in largest diameter). Both species are deepwater inhabitants being thus well differentiated on ecological grounds too, when compared to the new species apparent shallow, warm, mangrove restricted distribution. One further diagnostic character stems from C. schmidti s much stouter choanosomal oxeas.
Craniella cranium is a problematic species in need of revision. Originally described from the Boreal Eastern Atlantic, the species has subsequently been reported from several other, mostly Boreal/Temperate localities, on many distinct biogeographic provinces. On an exclusively biogeographical probability rationale, Boreal records on both the Atlantic and the Pacific are likely, Southern Temperate records and a postulated antitropical distribution are less likely, and Tropical records and their implied cosmopolitanism are much less likely. With this in mind, it is very improbable on biogeographical grounds alone that C. cranium will be found on warm, shallow Tropical Brazil. Consequently, Carters (1890) record of C. cranium for the Fernando de Noronha Arquipelago, despite not revised here, is most probably equivocal. We have taken Van Soest et alls (2000) description as a recent revision of Boreal Western Atlantic C. cranium , and used it for a comparison with C. quirimure sp.nov. The Boreal species has much stouter cortical (up to 55 m thick) as well as choanosomal oxeas (up to 70 m thick). Its triaenes, both pro and ana, were reported to be much longer than those in the new species (respectively, up to 8500ìm and 20000ìm). Sigmaspires in C. cranium may reach 20 m in diameter while in C. quirimure sp.nov. these are smaller than 15ìm. Given all these morphological differences, and keeping in mind both species apparent very low biogeographic relatedness, C. quirimure sp.nov. is considered very well differentiated from C. cranium .
The two remaining Craniella reported from the Tropical (or nearly so) Atlantic are C. insidiosa and C. laminaris . The first one was poorly described by Schmidt (1870) and was considered unrecognizable already by Sollas (1888). No further comments can be made here on it status. C. laminaris , in spite of originally described from Newport River (eastern U.S.A.), a locality nearly 9000 km away from Bahia, appears to be the morphologically closest to the new species. Both share the very shallow water habitat, as well as comparably large cortical oxeas and sigmaspires, and possess additional categories of protrianes (cf. George & Wilson, 1919). A series of traits can be used to separate them. Cortical oxeas can be much stouter (up to 28 m thick) in C. quirimure sp.nov., choanosomal oxeas can be much longer (up to 4551 m) and stouter (up to 28 m thick), and anatriaenes can be over 4x larger (up to 9600 m). The new species is thus considered well distinguished from other congeners in the Tropical west Atlantic.

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |