
Gymnura micrura ( Bloch & Schneider 1801 )
|
publication ID |
https://doi.org/10.11646/zootaxa.4332.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:C7D6FF64-5813-4AAB-AE40-C24ABB74433F |
|
DOI |
https://doi.org/10.5281/zenodo.6039806 |
|
persistent identifier |
https://treatment.plazi.org/id/03A787B2-FF98-FFD4-A2BE-8F15344DFDA9 |
|
treatment provided by |
Plazi |
|
scientific name |
Gymnura micrura ( Bloch & Schneider 1801 ) |
| status |
|
Gymnura micrura ( Bloch & Schneider 1801) View in CoL
Smooth (or Lesser) Butterfly Ray
( Figs. 1–10 View FIGURE 1 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 View FIGURE 7 View FIGURE 8 View FIGURE 9 View FIGURE 10 , 34a View FIGURE 34 , 35 View FIGURE 35 , 36a View FIGURE 36 – 38a View FIGURE 38 , 39–41 View FIGURE 39 View FIGURE 40 View FIGURE 41 , 42a View FIGURE 42 – 44a View FIGURE 44 , 45 View FIGURE 45 , 46a View FIGURE 46 – 48a View FIGURE 48 ; Tabs. 1–2)
Raja micrura Bloch & Schneider, 1801: 360 (original description, type locality: Suriname)
? Trygon micrura: Müller, 1837: 40 (not seen: Rio de Janeiro, according to Bigelow & Schroeder [1953] identification probable because said to agree with description of Raja micrura of Bloch & Schneider 1801).
Pteroplatea maclura: Puyo, 1936: 79 , 250 ( French Guiana; not seen).
Pteroplatea maclura (in part): Müller & Henle, 1841: 169 (description, specimens from Suriname and Brazil only); Duméril, 1865: 614 (description, specimens from Brazil only, size probably erroneous as the New York specimen [= G. lessae View in CoL , sp. nov.] recorded with a width of 2 meters is the same specimen analyzed by Müller & Henle, 1841, who had earlier characterized it as small); Günther, 1870: 487 (description, specimen from Brazil only).
Pteroplatea micrura: Metzelaar, 1919: 8 View in CoL , 199 (listed, Trinidad, perhaps Curaçao).
Pteroplatea micrura View in CoL (in part): Meek & Hildebrand, 1923: 87 (not present in Panamá, color description matching G. lessae View in CoL , sp. nov.).
Gymnura micrura: Nishida, 1990: 65 View in CoL , 66, 83, 84, fig. 59B (skeletal morphology, clasper;?Suriname); Lovejoy, 1996: 214, 215, 223, 224, 227, 228, 230, 236, 242, 256, fig 7E (lateral line canals, skeletal morphology; Guianas and Suriname); Mazzoleni & Schwingel, 1999: 144, 116 (listed, Southern Brazil); Menni & Stehmann, 2000: 92 (distribution, South America); Furtado Jr. et al. 2003: 8, 9 (fisheries, North Brazil); Carvalho et al., 2004: 10, 33, 38, 39, 74, 79, 82, 86, 87, figs. 15, 31, 35B, 36C-B, 38C, 39A-B (skeletal morphology, Suriname); Meneses et al. 2005: 80, 82 (listed; Sergipe, Brazil); Nunes et al. 2005: 51, 52, 53 (inventory; Maranhão, Brazil); Yokota & Lessa, 2006: 349 –360 (nursery area, Northeastern Brazil); Yokota & Lessa, 2007: 249 –257 (reproductive biology, Northeastern Brazil); Basílio et al. 2008: 67 – 69 (checklist; Curu River estuary, Ceará, Brazil); Lessa et al. 2008 (fisheries, Northeastern Brazil); Basilio et al. 2009: 41 (listed; Ceará, Brazil); Bornatowski et al. 2009: 3 (listed; Southern Brazil); Nunes & Piorski, 2009: 479 –482 (presence dorsal fin; Maranhão, Brazil); Reis et al. 2012: 217 –219 (albinism; Alagoas, Brazil); Yokota et al., 2012: 1315 –1326 (reproductive biology, Northeastern Brazil); Ragno, 2013: 57, 58, 130, 177, fig. 46 (lateral line; Rio Grande do Norte, Brazil); Yokota et al., 2013: 1325 –1329 (diet, Northeastern Brazil); Fontenelle & Carvalho, 2015: 7 –9, figs. 8, 9E, 10E (brain morphology; Rio Grande do Norte, Brazil); Garcia Jr. et al., 2015: 4 (checklist; Rio Grande do Norte, Brazil); Willems et al., 2016: 36, 38, 40, 41 (fisheries, Suriname).
Gymnura micrura View in CoL (in part): Rosenberger, 2001: 615, 616, 618, 620, 621, 623–627 (systematics, specimen FMNH 89993 only [Suriname],?FMNH 89992); McEachran & Carvalho, 2002: 577 (identification guide, Western Central Atlantic; illustration is of G. lessae View in CoL , sp. nov.).
Not Raja maclura: Lesueur, 1817: 41 , pl. not numbered (original description, Newport, Rhode Island, junior synonym of G. altavela View in CoL ).
Not Pteroplatea micrura: Müller & Henle, 1841: 169 View in CoL (Java, India, Red Sea; = Gymnura poecilura View in CoL [ Shaw, 1804]); Cantor, 1849: 1409 (= G. poecilura View in CoL [ Shaw, 1804]); Blyth, 1860: 29 (= G. poecilura View in CoL [ Shaw, 1804]); Day, 1865: 278 (= G. poecilura View in CoL [ Shaw, 1804]); Duméril, 1865: 613 (description, mouth of Ganges, = G. poecilura View in CoL , [ Shaw, 1804]); Günther, 1870: 487 (= G. poecilura View in CoL [ Shaw, 1804]); Day, 1878: pl. CXCIV, fig. 2 (= G. poecilura View in CoL [ Shaw, 1804]); Day, 1889: 56, fig. 23 (= G. poecilura View in CoL [ Shaw, 1804]); Wood-Mason & Alcock, 1891: 359 –367, pl. 7–8 (embryonic development, = G. poecilura View in CoL [ Shaw, 1804]); Alcock, 1892: 1 –8, pl. 4 (embryonic development, = G. poecilura View in CoL [ Shaw, 1804]); Annandale, 1909: 39 (= G. poecilura View in CoL [ Shaw, 1804]); Pillay, 1929: 353 (= G. poecilura View in CoL [ Shaw, 1804]).
?Not Gymnura View in CoL sp. cf. micrura: Compagno & Last, 1999: 1507, 1509 (identification guide, Western Central Pacific); Randall & Lim, 2000: 583 (checklist, South China Sea); Jacobsen & Bennett, 2009: 1, 21, 22, 24, 25 (taxonomic revision, Indo-West Pacific).
Not Gymnura View in CoL micrura: Fowler, 1941: 455 (description based on a G. australis View in CoL [Ramsay & Ogilby] specimen [USNM 39978]); Talwar & Kacker, 1984: 181 (= Gymnura poecilura View in CoL [ Shaw, 1804]); Raje, 2003: 89, 91–95 (probably equals Gymnura poecilura View in CoL [ Shaw, 1804]); Raje et al. 2007 (= Gymnura poecilura View in CoL [ Shaw, 1804]).
Neotype. USNM 156714 ( 289 mm DW, adult male), Suriname, 6o20’45”N, 54o56’30”W, 0 to 26m, Vessel Coquette, station 145, trawl, 30.v.1957, accession num. 215120, USNM 440341 recatalogued from USNM 156714 ( Fig. 1 View FIGURE 1 ). Designated herein.
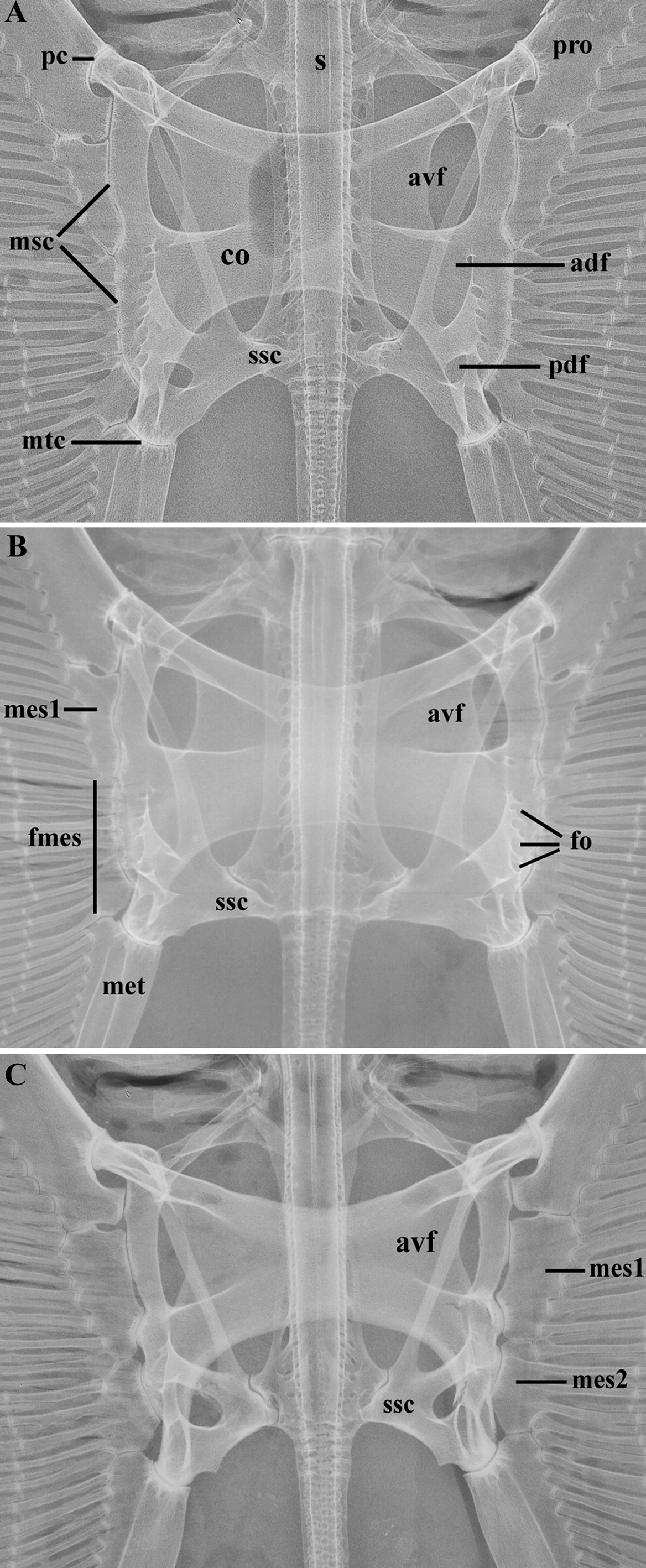
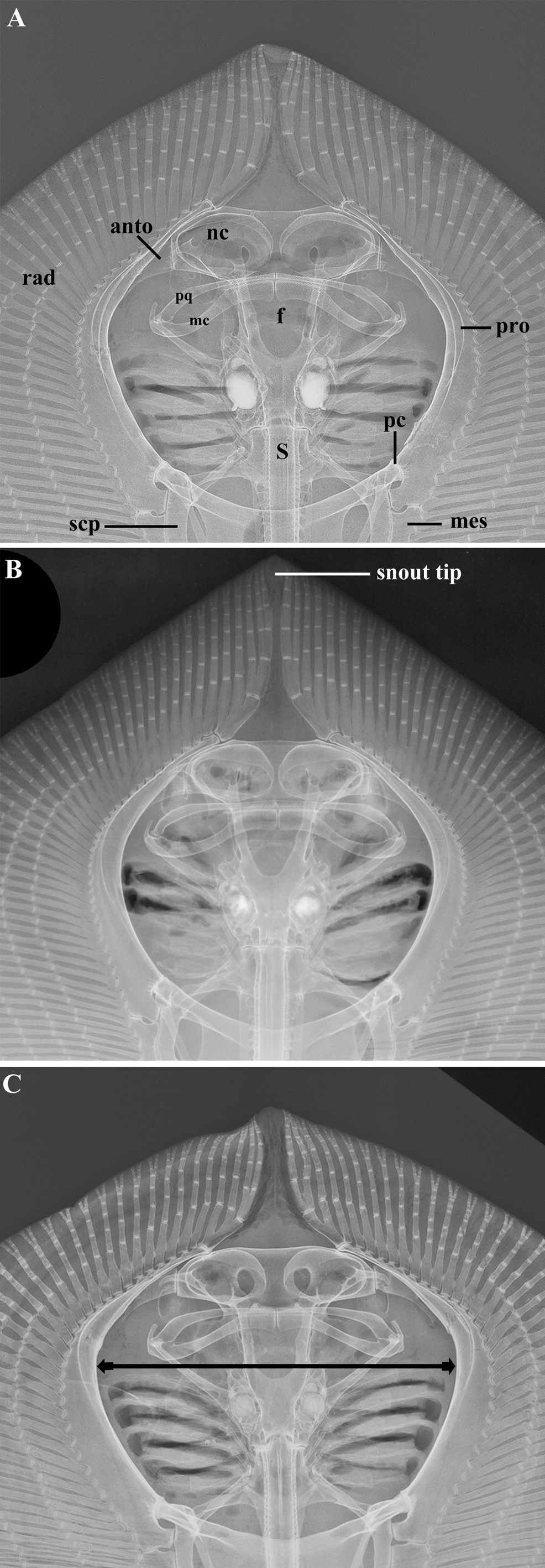
Diagnosis. A small to medium size butterfly ray occurring in the Western South Atlantic distinguished from congeners (except the two new species described in this study) by the combination of the following characters: absence of spiracular tentacle and caudal stings; dorsal fin usually absent (a small dorsal fin may be present in some males); tail relatively short (mean post-cloacal length 22% DW) and prominently banded, presenting 3 to 5 black bands that may be less distinct in large adults. Gymnura micrura is distinguished from G. lessae , sp. nov. and G. sereti , sp. nov. by its usually uniformly brown or gray dorsal disc, without any vermiculate pattern ( vs. dorsal side brownish, usually with a vermiculate pattern in both new species). Gymnura micrura may be further distinguished from G. sereti , sp. nov. by the following characters: clasper of mature males more slender and longer, Lclasper 9.3–11% DW ( vs. clasper stouter and shorter, Lclasper 6.8–9.2% DW); cranial fontanelle U-shaped ( vs. keyhole-shaped fontanelle); mesopterygium divided into one anterior solid element and 5–7 smaller fragments ( vs. mesopterygium divided into two solid elements); distance between anteroventral (AVF) and posteroventral (PVF) fenestrae of scapulocoracoid representing 30–35% of scapulocoracoid length ( vs. distance between AVF and PVF representing about 20% of scapulocoracoid length); lateral projection of the base of synarcual starting at its anterior third ( vs. lateral projection of the base of synarcual starting synarcual half-length); thicker proximal portion of Meckel’s cartilage representing 28–40% of Meckel’s cartilage width ( vs. 10–24%); lower number of diplospondylous vertebrae (mean 98 vs. 110, respectively), and higher number of radials associated to the mesopterygial fragments compared to the second mesopterygial element of G. sereti (range 15–17 vs. 10–15, respectively). Gymnura micrura is most similar to G. lessae , sp. nov., but may be further differentiated from it by the following characters: dorsal contour of hyomandibula with two conspicuous humps ( vs. dorsal contour of the hyomandibula with a conspicuous proximal protuberance followed by an inconspicuous distal protuberance); projection of the anteroventral fenestra of coracoid bar in the form of a funnel with a large opening and closing ( vs. projection of the anteroventral fenestra with a relatively narrower opening and end) and lower number of pectoral radials (range 112–119 vs. 118–127, respectively). Other diagnostic differences are described below.
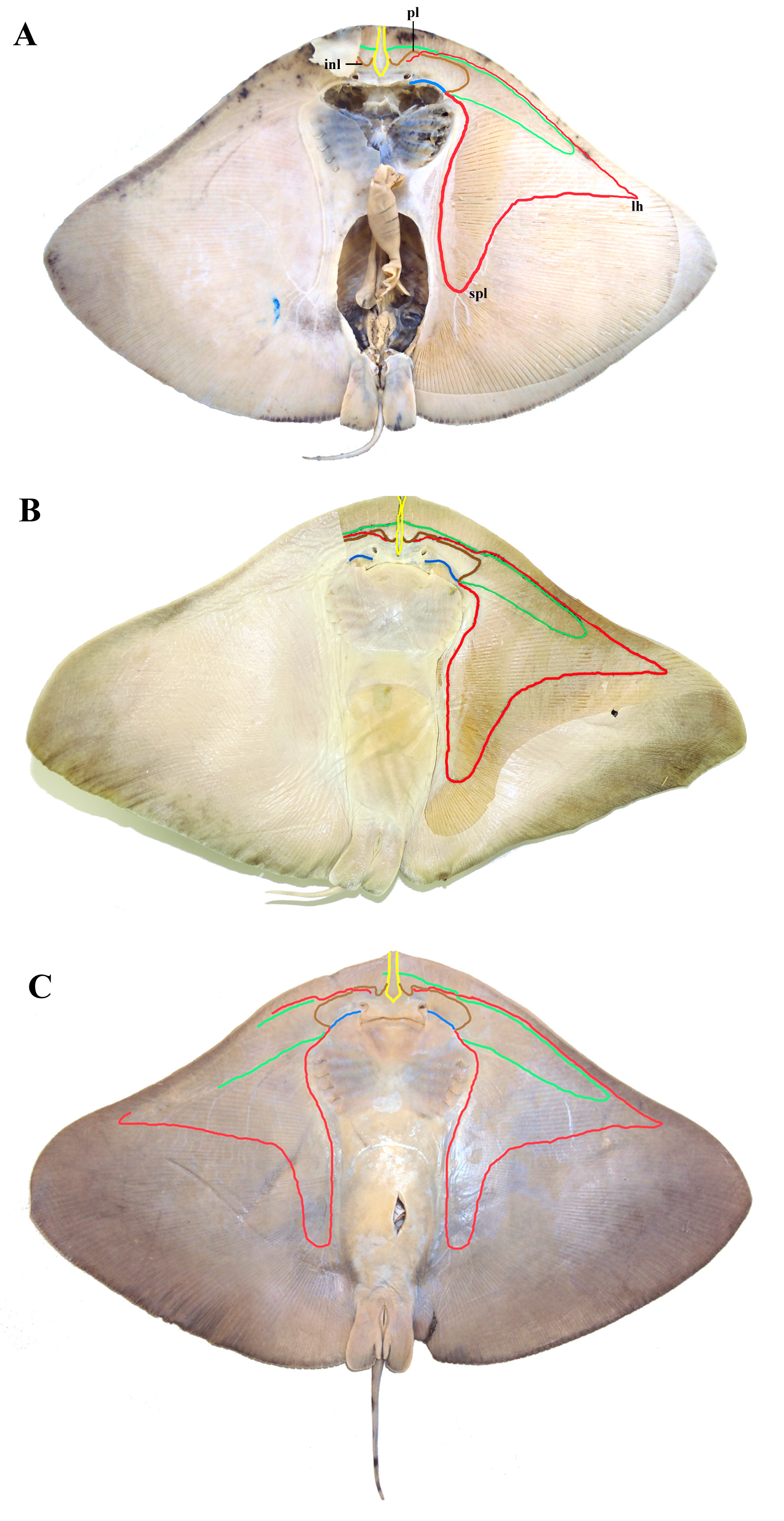
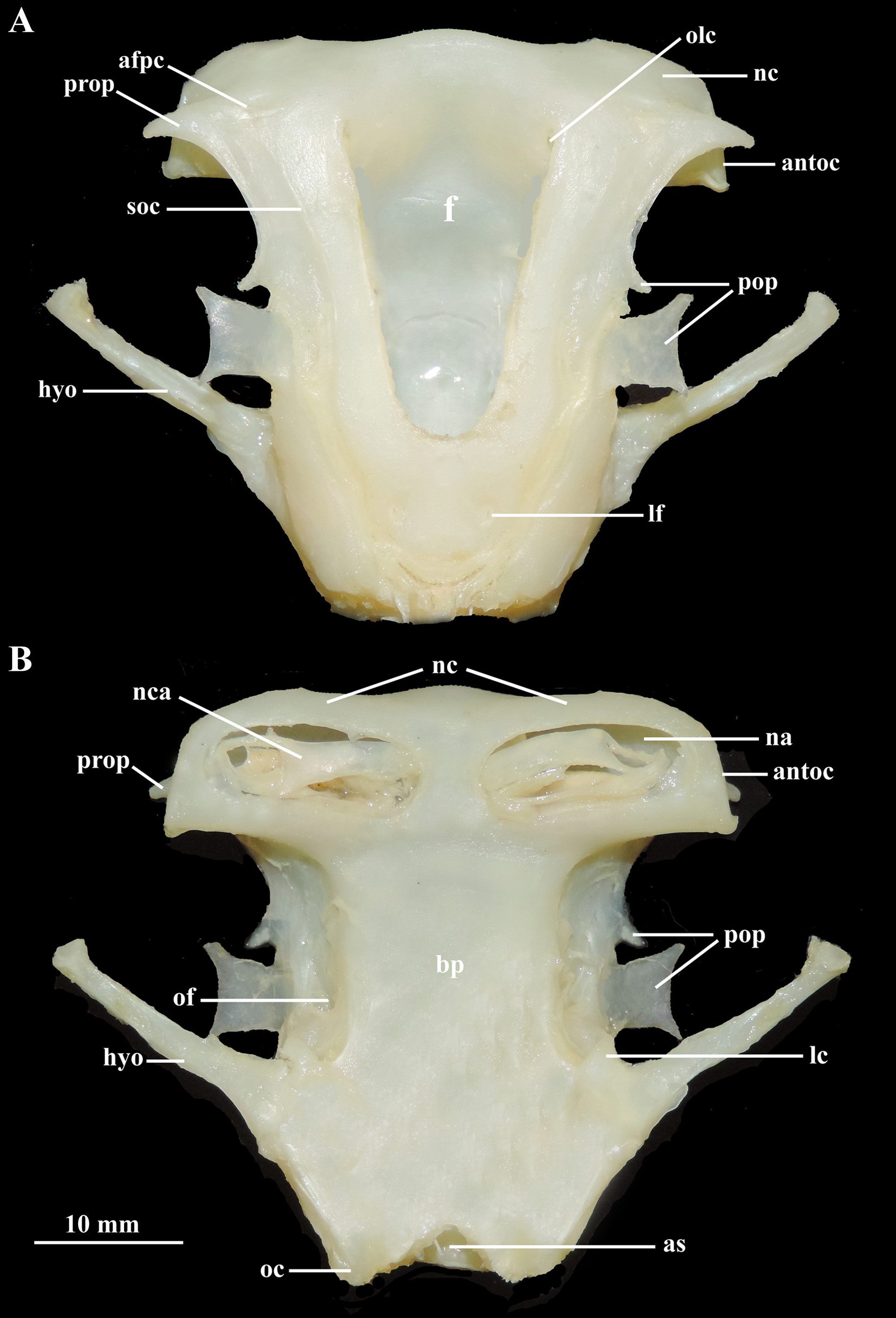
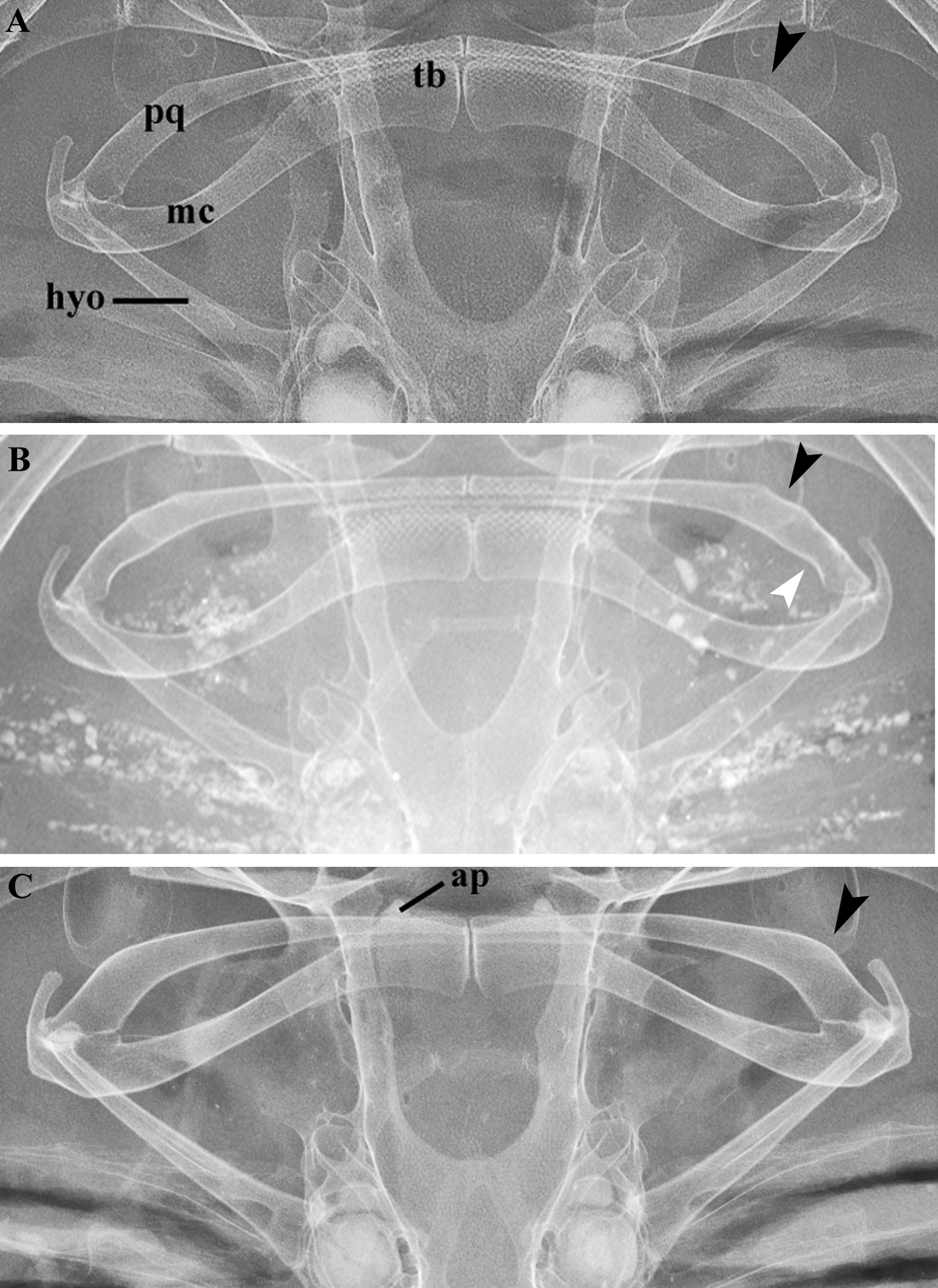
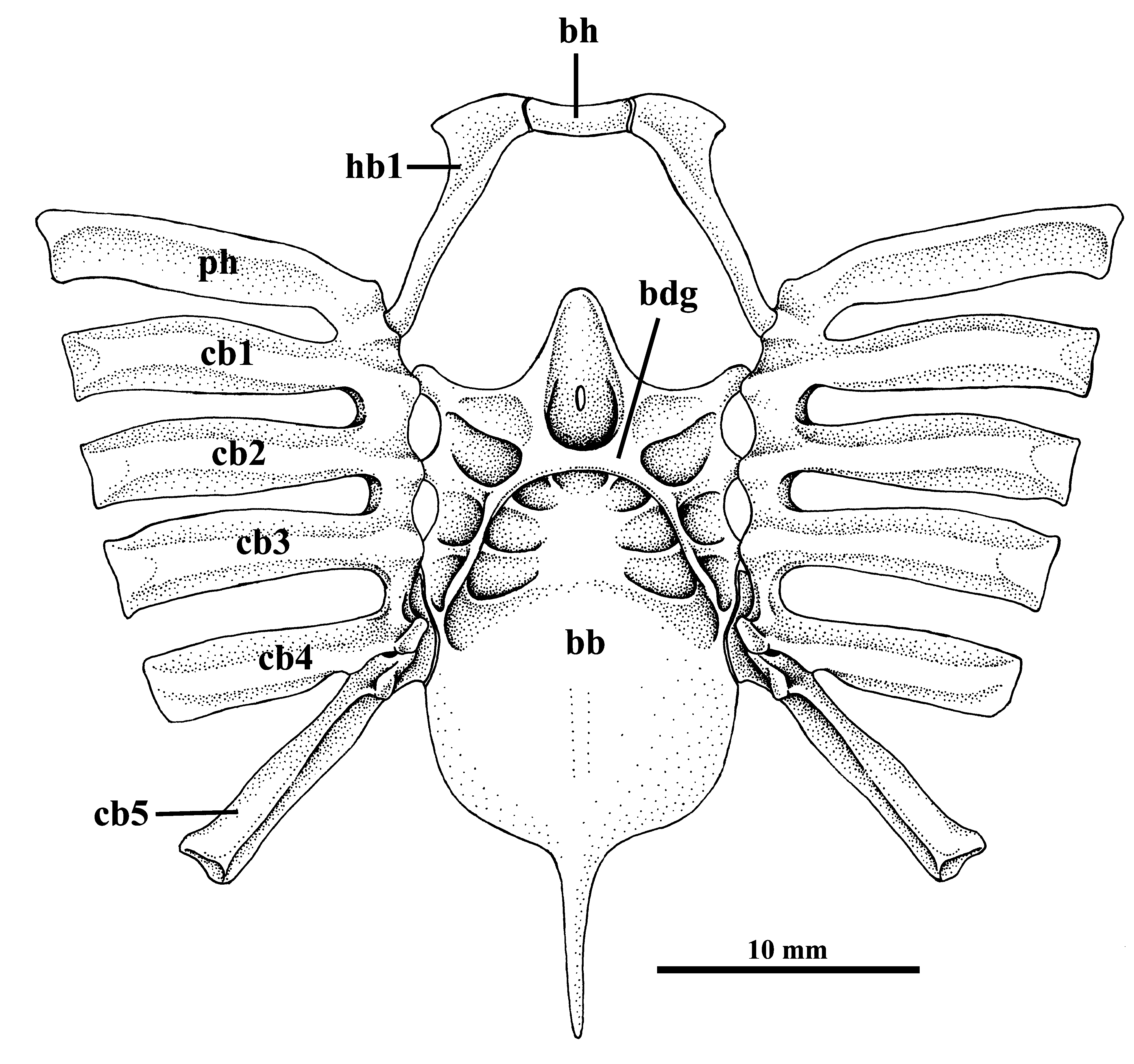
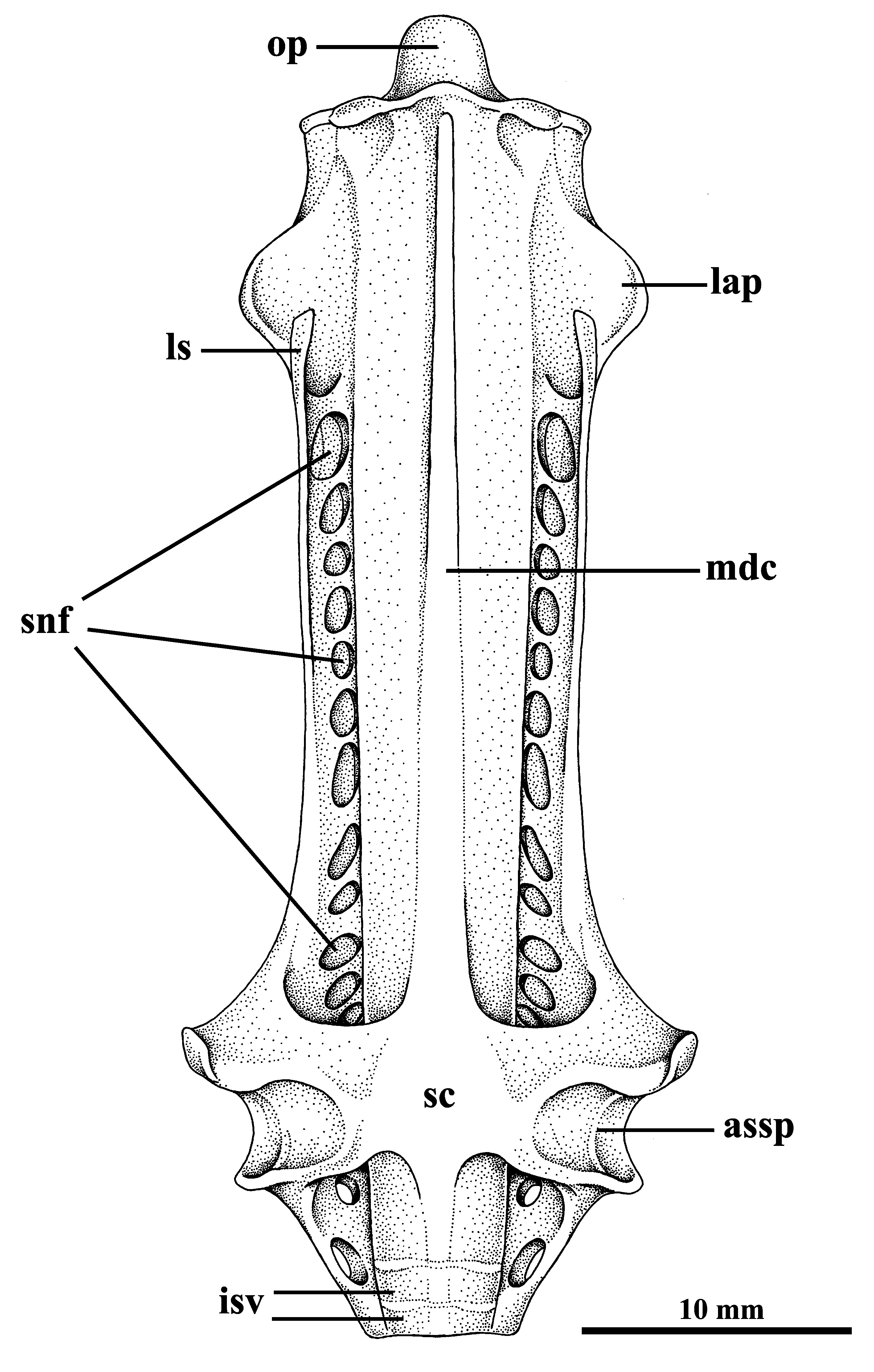
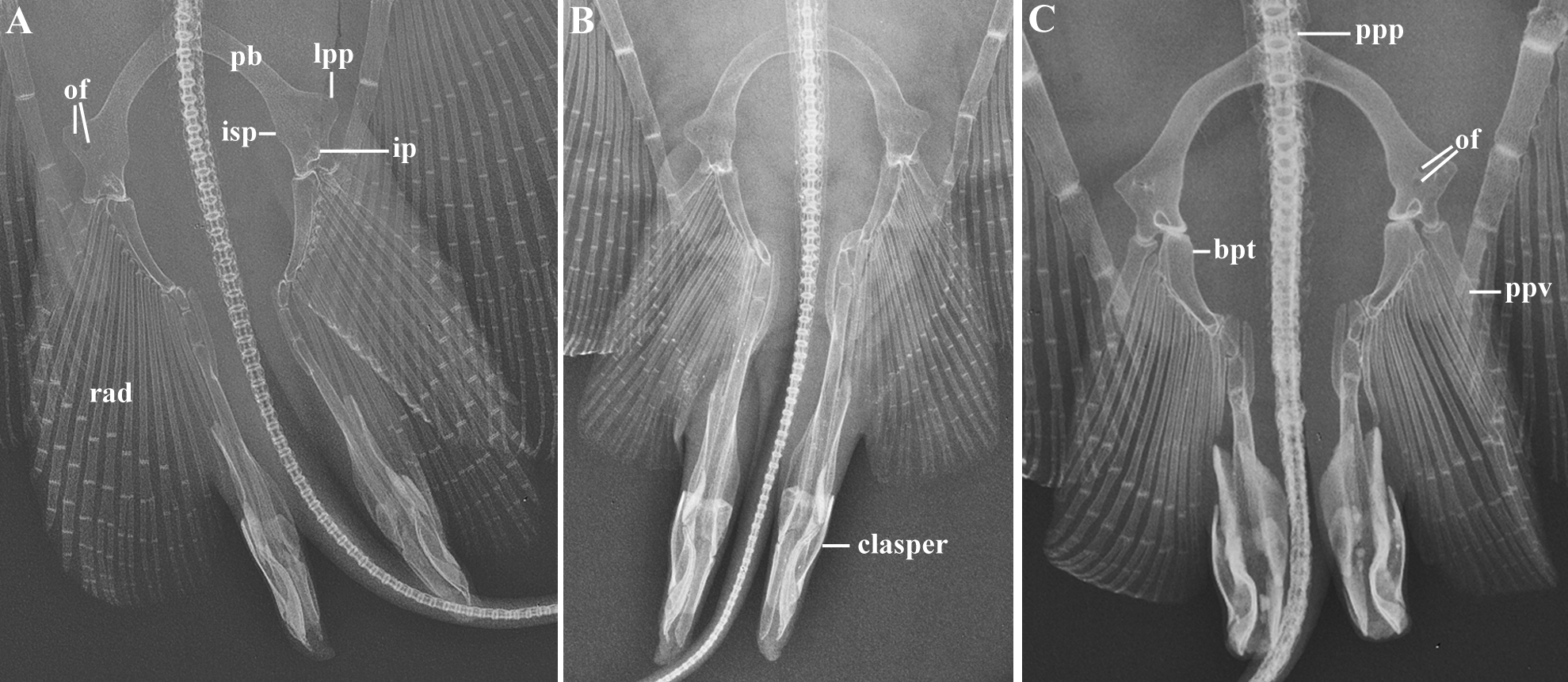
Description. Measurements are presented in Table 1; meristic data in Table 2. The following description is based on all specimens examined. Throughout the text the proportions are presented as: minimum value–maximum value (mean). See Figs. 1–4 View FIGURE 1 View FIGURE 2 View FIGURE 3 View FIGURE 4 , 5a–h View FIGURE 5 , 6a View FIGURE 6 and 7–9 View FIGURE 7 View FIGURE 8 View FIGURE 9 for external morphology and color, Fig. 34a View FIGURE 34 for ventral lateral-line pattern, Figs. 5i View FIGURE 5 , 6b View FIGURE 6 , 35 View FIGURE 35 , 36a View FIGURE 36 – 38a View FIGURE 38 , 39–41 View FIGURE 39 View FIGURE 40 View FIGURE 41 , 42a View FIGURE 42 – 44a View FIGURE 44 , 45 View FIGURE 45 and 46a View FIGURE 46 – 48a View FIGURE 48 for skeletal morphology and Fig. 10 View FIGURE 10 for geographical distribution.
External morphology. Disc lozenge shaped, 1.54–1.92 (1.75) times broader than long [1.54–1.72 (1.64) for adult males; 1.75–1.92 (1.81) for adult females]. Trunk strongly flattened, slightly raised above scapular region and posterior head. Snout relatively short and obtuse with a subtle lobe at snout tip. Sexual dimorphism evident in disc shape, mainly between adults, with adult males presenting proportionally longer snouts with smaller angles, which confers a triangular shape to the anterior half of the disc ( Fig. 2 View FIGURE 2 ). Preoral snout length 8.6–14 (11)% DW [8–11 (9)% DW in adult females; 12–14 (13)% DW in adult males], preorbital snout length 6.1–13 (9.4)% DW [7–10 (8)% DW in adult females; 10–13 (11)% DW in adult males], preorbital snout width 31–40 (37)% DW, postspiracle snout width 42–54 (49)% DW. Anterior margins of disc with a slight medial concavity and becoming weakly convex towards extremities (in adult males these curves may be less noticeable, with the anterior margins almost straight in some males) ( Fig. 2 View FIGURE 2 ); pectoral-fin apices acutely angular (sometimes moderately angular in adult males); posterior margins weakly convex; free rear tip broadly rounded; axis of greatest width positioned posteriorly to one-half disc length. Pelvic fin single lobed, rectangular, their corners rounded ( Fig. 3 View FIGURE 3 ). Skin entirely smooth, without denticles on dorsal and ventral surface of the disc.
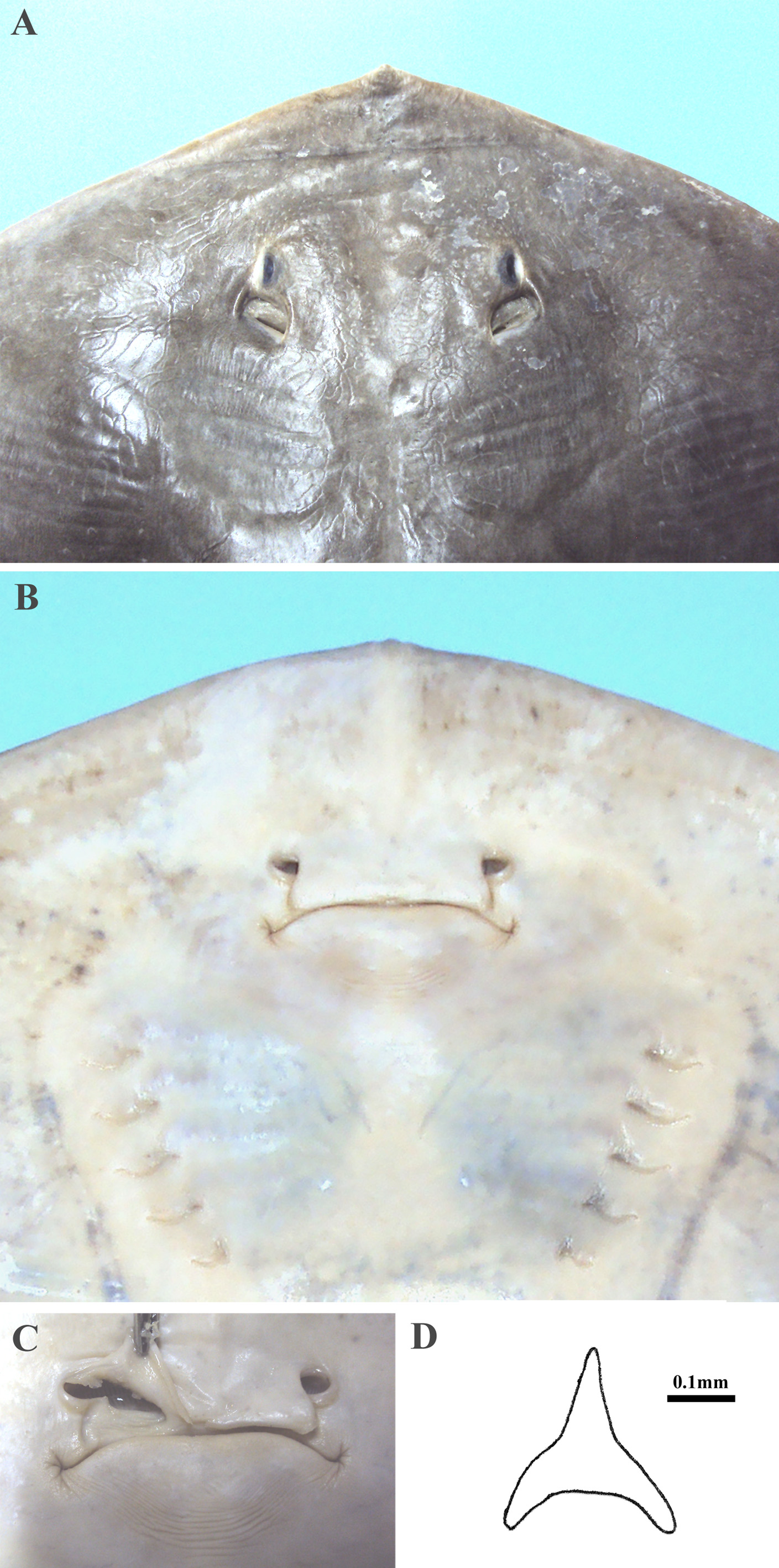
Interorbital space broad, interorbital width 7.9–9.8 (8.7)% DW. Eyes dorsolateral, small, oval and protruding slightly; eye diameter 1.2–2.2 (1.7)% DW, representing 13–25 (19)% of interorbital width; orbit diameter 1.9–3.5 (2.5)% DW, representing 59–108 (86)% of spiracle length. Eyes more protruded and relatively larger in embryos. Spiracles immediately following the eyes, relatively large, lozenge-shaped, contour of the inner spiracular margin concave ( Fig. 4a View FIGURE 4 ); spiracle length 2.1–4 (2.9)% DW, 1.1–2.7 (1.8) times eye diameter; its inner posterior margin without tentacles.
Ventral head length 20–27 (23)% DW. Nostrils narrowly oval, diagonally directed, only the circular distal margin not covered by nasal curtain, posterior lateral margin with lobe ( Fig. 4c View FIGURE 4 ); nostril length 1.8–4.3 (3.2)% DW [2.5–3.4 (2.9)% DW in adult females; 3.1–4.3 (3.7)% DW in adult males], 1.0–3.0 (1.8) times internasal width; internasal width 1.1–2.7 (1.8)% DW. Anterior nasal flaps medially expanded and fused into a broad, skirt-shaped, posteriorly expanded nasal curtain that covers the internasal space and reaches mouth ( Fig. 4b View FIGURE 4 ). Nasal curtain skirtshaped, weakly to moderately bilobed, wide, its width 1.9–4.3 (3.0) times length; lateral margin concave; posterolateral apices rounded; posterior margin straight to weakly concave, a small gap may be present medially.
Mouth relatively wide, its width 7.5–10 (8.5)% DW, 32–45 (38)% head length, 0.9–1.8 (1.2) times nasal curtain width; without papillae on floor or labial folds, although some striations may originate radially from mouth corners; lower lip arched rearward toward corners, uniformly convex or weakly to moderately concave along medial region, without any lump or knob ( Fig. 4b View FIGURE 4 ); a strip of corrugated skin forms a half-circle below mouth; strips ends at the mouth corners. Small, numerous and closely crowded teeth in bands; teeth with one medial, pointed cusp directed towards inside of mouth; tooth base somewhat swollen, entirely twisted to the opposite direction; in labial face, tooth resembling a three-armed boomerang ( Fig. 4d View FIGURE 4 ); teeth similar between jaws and sexes; their number increasing with growth.
Gill slits moderately S-shaped, first four gill slits markedly larger than fifth ( Fig. 4b View FIGURE 4 ). Distance between gill slits decreasing posteriorly; distance between inner ends of fifth pair 60–74 (67)% distance between inner ends of first pair; distance between inner ends of first pair 15–18 (16)% DW, 58–82 (72)% ventral head length, 6.3–14.0 (9.3) times internasal width; distance between inner ends of fifth pair 10–12 (11)% DW, 40–56 (48)% ventral head length, 4.1–9.3 (6.3) times internasal width.
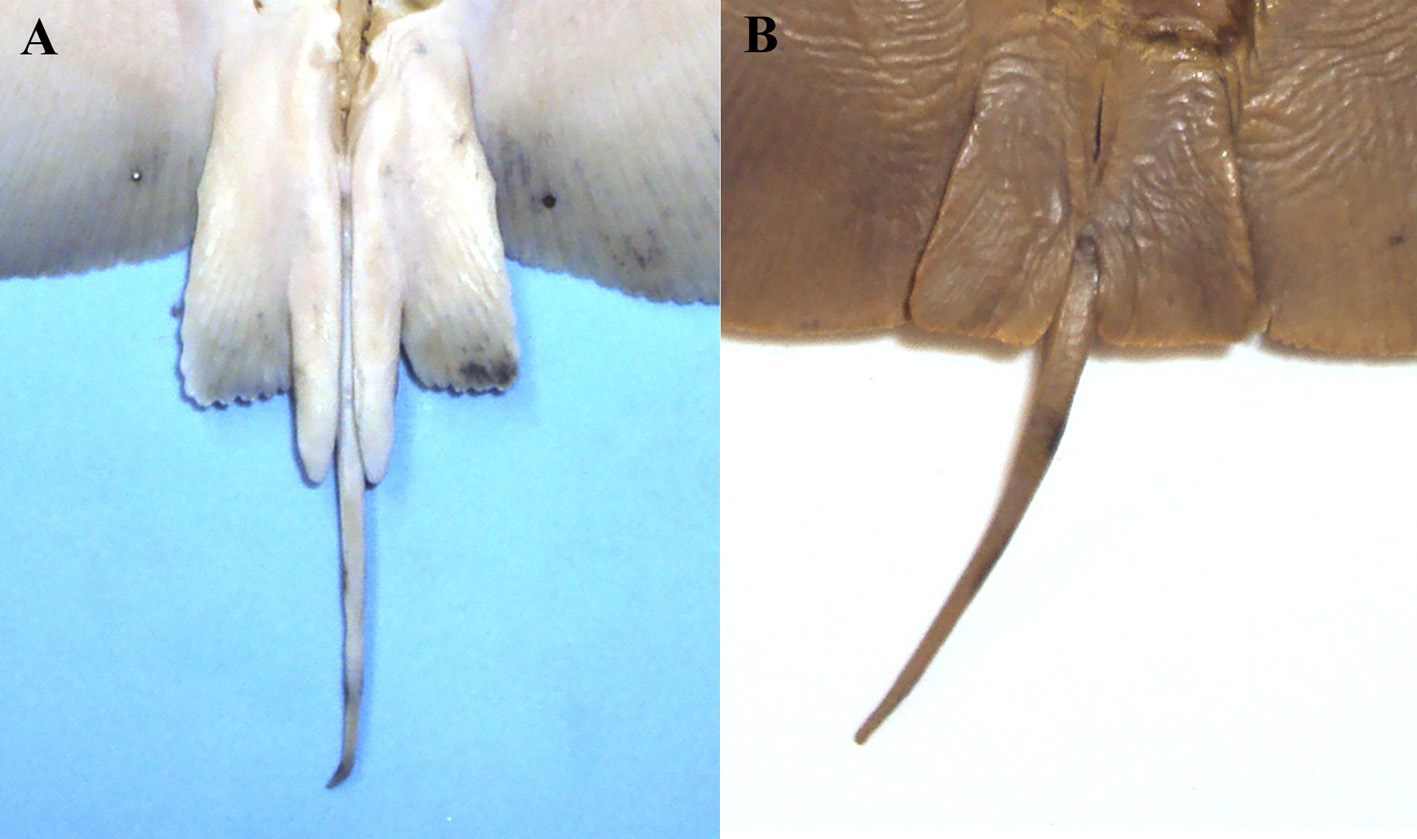
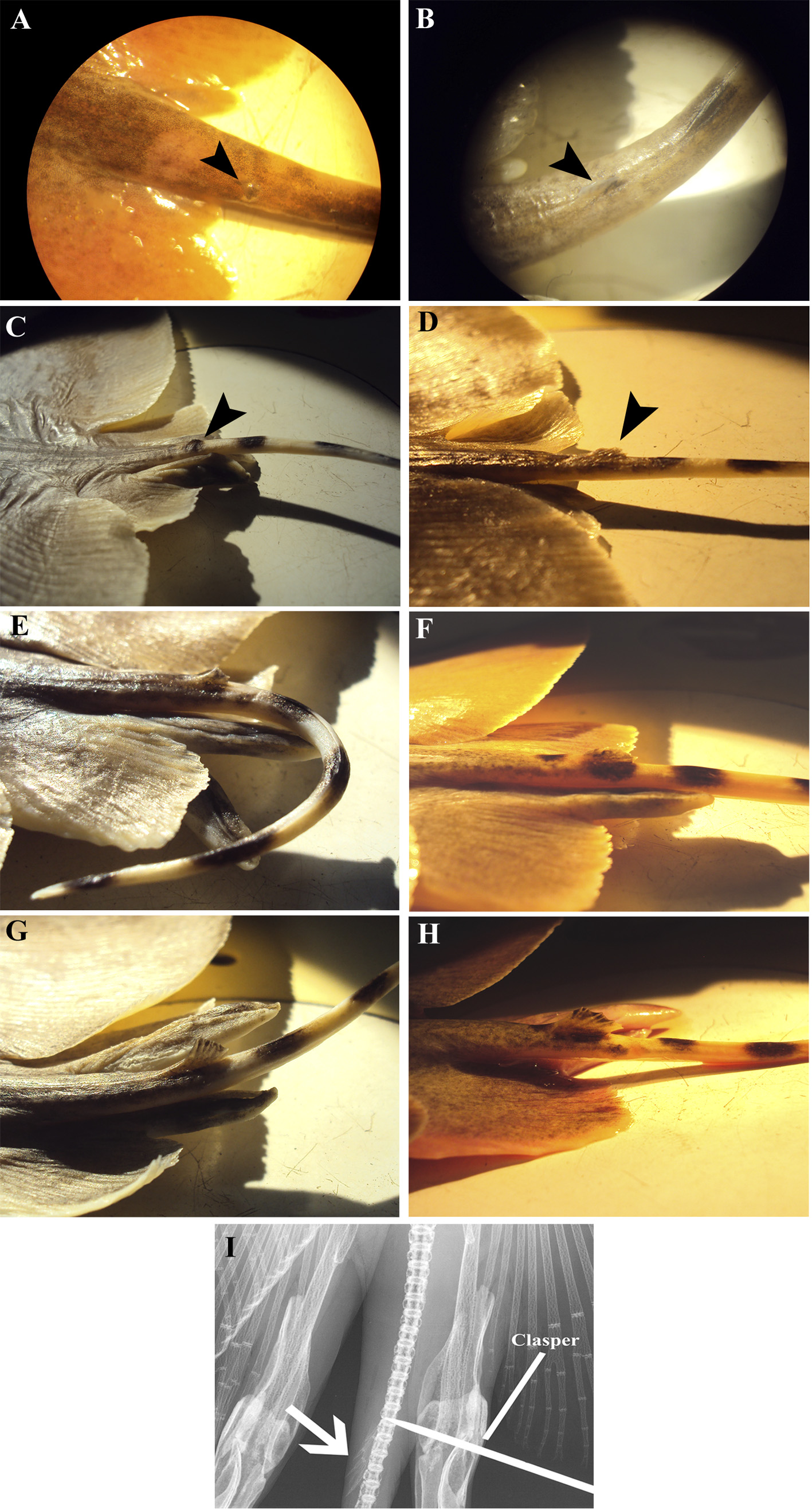
Tail slender and short, whip-like, cross-banded (see section on color), tapering toward tip, without caudal stings at any age ( Fig. 9 View FIGURE 9 ); postcloacal tail 17–30 (22)% DW, 30–47 (38)% DL, 30–56 (45)% precloacal length; tail usually lacking dorsal fin, however some males (about 25% of males) may present a small plesodic dorsal fin at tail base with varying degrees of development ( Figs. 5a–h View FIGURE 5 ); when present the dorsal fin is situated at the level of free rear tips of pelvic fin, distance from snout tip to anterior base of the dorsal fin representing 76–80 (78)% of total length; low longitudinal skin folds present on dorsal and ventral surfaces of tail, skin folds not reaching the tail tip.
Clasper of mature males cylindrical, somewhat depressed, relatively slender and longer (compared to G. sereti , sp. nov.) ( Fig. 6 View FIGURE 6 ); distance from posterior margin of cloaca to clasper tip 9.3–11.0 (9.9)% DW; clasper tapering slightly distally; clasper tip conical, bluntly pointed to pointed, not calcified; portion not calcified relatively shorter than in G. lessae , sp. nov. and G. sereti , sp. nov.; apopyle (on the anterior dorsal surface) connected to hypopyle by a long, dorsomedial, posteriorly curved clasper groove; spermatic duct almost reaching clasper tip; rhipidion and pseudorhipidion (“small flap” of Nishida 1990) absent; a long ventral pseudosiphon (“SAC 1” of Nishida 1990) laterodistally situated to hypopyle; a well-developed dorsal pseudosiphon (“SAC 2” of Nishida 1990) on inner margin of clasper; dorsal pseudosiphon more posteriorly located when compared to G. sereti , sp. nov.; ventral surface of clasper entirely smooth.
Coloration. Dorsal side usually uniformly light to dark brown, grayish, sometimes slightly roseate, not vermiculated ( Figs. 7a View FIGURE 7 , 8 View FIGURE 8 ); irregular dark patches or irregular dark round spots may be present, especially in freshly caught specimens ( Figs. 7b, c View FIGURE 7 ). Living specimens may be variable in color according to the environment ( Figs. 7d, e View FIGURE 7 ), but they tend to become uniformly brown or gray when preserved. Ventral surface whitish to brownish, creamy, slightly roseate, yellowish or copper, generally darkening toward edges. Tail prominently cross-banded, with 3–5 black bands ( Fig. 9a View FIGURE 9 ); bands diffuse or less distinct in large specimens ( Fig. 9b View FIGURE 9 ).
Size, reproduction, diet and habitat. Size at birth ranging from 135 to 185 mm DW; size at 50% maturity was estimated at 269 (265–273) mm and 405 (401–409) mm for males and females, respectively ( Yokota et al. 2012). The largest male analyzed was 360 mm DW (MZUSP 9926, from Espírito Santo, Brazil), and males of this species shall reach a maximum of 450 mm DW. The largest female analyzed was 629 mm DW (sampled in Rio Grande do Norte, Brazil, by the first author) and the females reach around 800 mm DW. This species is matrotrophic viviparous (lipid histotrophy) and appears to be reproductively active throughout the year, producing 1–6 young per litter ( Yokota et al. 2012). Diet consists mainly of bony fishes with evidence of feeding specialization, ingesting intermittently proportionally larger fishes ( Yokota et al. 2013). This is a demersal species living along the coast on sandy and muddy bottoms, but also occurs in estuaries.
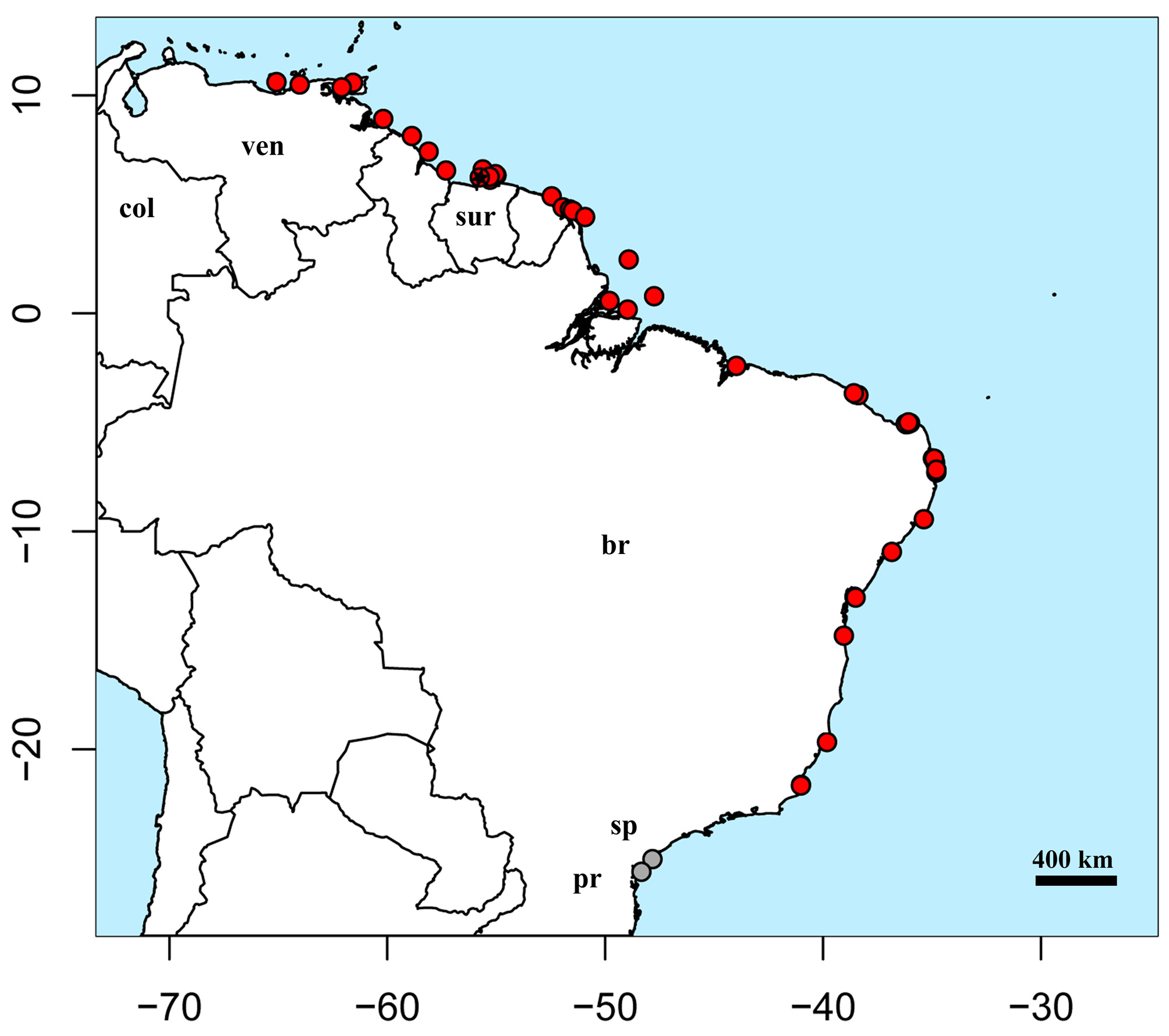
Geographical distribution. According to material examined, G. micrura is distributed in the Atlantic coast of South America from Venezuela (including Trinidad & Tobago) to Rio de Janeiro State, Brazil ( Fig. 10 View FIGURE 10 ). The species is also registered in São Paulo and Paraná States, Brazil (Gonzalez 1995, P. Charvet, pers. comm.). The species is relatively common in the north of South America and northern and northeastern regions of Brazil, with its abundance declining towards the south, where its occurrence may be related to seasonal warmer water. There are no confirmed records of this species in Colombia and Panama ( Meek & Hildebrand 1923, Mejía-Falla et al. 2007), and it is not known if the species occurs in the Caribbean Sea, beyond the Venezuelan coast.
Remarks. The original description of Gymnura micrura (as Raja micrura Bloch & Schneider 1801 ) is brief, nondiagnostic, presenting characters that are commonly shared by all species in the genus, and is not illustrated; however, the type locality is clearly stated as Suriname (“Habitat in Surinamo”, p. 360). This fixed the name micrura to the butterfly ray without caudal stings and spiracular tentacles commonly found in the region, in contrast to G. altavela ( Linnaeus 1758) , which possesses spiracular tentacles and caudal stings and does not occur in the area. Bloch & Schneider mention that type material would probably be housed in the “Museo Vaillanti Parisiis”. After an exhaustive search in the Muséum national d'Histoire naturelle in Paris and other collections, we are not able to locate any type material for G. micrura . At the same time, this type material was never mentioned in any study subsequent to the original description (e.g. Séret & McEachran 1986) and is considered to be unknown in Eschmeyer et al. (2017); the type material was probably lost or destroyed. As this species is central to a complex taxonomic problem, in which three distinct species were being identified under the name G. micrura (two of these are described as new here), the designation of a neotype for this species is expressly required to define the nominal taxon objectively and clarify its taxonomic status. The designed neotype matches the available information from the original type locality.
Fowler (1941) characterized the type locality of Bloch & Schneider as “likely erroneous” but without further comment. This author probably made this comment because the species he named G. micrura is not that of Bloch & Schneider, but a misidentification of Gymnura from the Indian Ocean and Indo-Pacific region. His description of G. micrura , for example, is based on a specimen of G. australis (Ramsey & Ogilby 1886) (USNM 39978). Hence, a type locality in South America for a species from the Indo-Pacific region is erroneous.
Müller & Henle (1841) mistakenly applied the specific name micrura to G. poecilura of Shaw (1804), and were followed by many others authors (e.g. Cantor 1849, Blyth 1860, Day 1865, Duméril 1864, Günther 1870, Day 1878, 1889, Wood-Mason & Alcock 1891, Alcock 1892, Annandale 1909, Pillay 1929). Although G. micrura and G. poecilura are well-separated species, this nomenclatural confusion lasts to the present (e.g. Talwar & Kacker 1984, Raje 2003, Raje et al. 2007) and it is common to find representatives of G. poecilura in fish collections worldwide identified as G. micrura (LY, pers. obs.). However, the relationship between G. micrura and an Indo- Pacific butterfly ray that lacks spiracular tentacles and has a short tail without caudal stings, which has been identified as G. cf. micrura (e.g. Compagno & Last 1999, Randall & Lim 2000, Jacobsen & Bennett 2009), needs to be resolved. This issue is being addressed in the revision of Indo-Pacific butterfly rays (Yokota & Carvalho, in prep).
Material examined. (143 specimens). Venezuela (18 specimens): ANSP 120086 About ANSP ( 12 specimens: juvenile female, 185 mm DW; juvenile female, 217 mm DW; juvenile female, 258 mm DW; juvenile male, 267 mm DW; juvenile male, 249 mm DW; juvenile male, 249 mm DW; adult female, 520 mm DW; adult female, 547 mm DW; adult male, 324 mm DW; adult male, 318 mm DW; juvenile female, 239 mm DW; juvenile male, 266 mm DW), Estado Sucre, Peninsula de Araya or Golfo de Cariaco ; ANSP 120162 ( 3 specimens: juvenile male, 200 mm DW; juvenile male, 199 mm DW; juvenile female, 267 mm DW), Estado Sucre, Caribbean Sea, Peninsula de Araya between Punta Homo and Punta Cardon ; USNM 222908 About USNM ( 3 specimens: juvenile female, 297 mm DW; juvenile male, 209 mm DW; juvenile female, 250 mm DW), Mouth of Rio Orinoco , 0 8o 56’N, 60o11’W GoogleMaps . Trinidad and Tobago ( 3 specimens) : BMNH 1931.12 .5.117 (juvenile female, 222 mm DW), Port Spain, fish Market ; BMNH 1932.2.8.3-4 ( 2 specimens: juvenile male, 260 mm DW; juvenile male, 269 mm DW), Gulf of Paria . Guiana ( 6 specimens) : BMNH 1961.8.31.34-36 (3 specimens: juvenile male, 227 mm DW; juvenile male, 258 mm DW; adult male, 269 mm DW), 0 7o26’N, 58o06’W; BMNH 1961.8.31.37-38 (2 specimens: adult male, 312 mm DW; female, 413 mm DW), 0 6o34’N, 57o18’W; ZMB 34703 (juvenile female, 202 mm DW). Suriname (17 specimens): RMNH D.2476 (dried); USNM 440341 (8 specimens: juvenile female, 251 mm DW; juvenile female, 254 mm DW; juvenile female, 277 mm DW; juvenile female, 306 mm DW; juvenile female, 335 mm DW; juvenile male, 205 mm DW; juvenile male, 225 mm DW; juvenile male, 235 mm DW), 0 6o21’N, 54o59’W; USNM 156716 (4 specimens: juvenile female, 188 mm DW; juvenile female, 225 mm DW; juvenile female, 280 mm DW; juvenile male, 271 mm DW), 0 6o24N, 55o02W; USNM 205354 (female, 488 mm DW), 0 6o37’N, 55o37’W; USNM 222601 (subadult male, 268 mm DW), 0 6o17’N, 55o18’W; ZMA 109.539 (adult male, 345 mm DW), 0 6o15’N, 55o45’W. French Guiana ( 10 specimens): CAS 50886 (juvenile male, 207 mm DW), 0 5o 23’N, 52o27’W GoogleMaps ; MNHN 2445 (juvenile female, 315 mm DW); USNM 222598 (3 specimens: juvenile female, 245 mm DW; juvenile female, 271 mm DW; subadult male, 282 mm DW), 0 4o47’N, 51o37’W; USNM 222615 (juvenile male, 235 mm DW), 0 4o43’N, 51o29’W; USNM 222616 (3 specimens: juvenile female, 270 mm DW; juvenile female, 292 mm DW; juvenile male, 247 mm DW), 0 4o26’N, 50o55’W; USNM 222622 (adult male, 323 mm DW), 0 4o26’N, 50o55’W. Amapá, Brazil (1 specimen): USNM 156822, 0 2o29’N, 48o55’W. Pará, Brazil (5 specimens): USNM 222597 About USNM ( 5 specimens: female, 377 mm DW; juvenile male, 187 mm DW; subadult male, 281 mm DW; subadult male, 292 mm DW; female, 324 mm DW), off mouth of Amazon River , 0 0o 48’N, 47o45’W. Maranhão, Brazil ( 1 specimen) GoogleMaps : MZUSP 72827 View Materials (female, 362 mm DW), São Luís, Rio Curuca. Ceará, Brazil ( 7 specimens) : SU 51597 ( 7 specimens: 4 embryos, 140 to 150 mm DW; embryo female, 185 mm DW; juvenile male, 197 mm DW; juvenile male, 250 mm DW), Fortaleza, Mucuripe, 0 3o 43’00”S, 38o28’59”W. Rio Grande do Norte, Brazil ( 52 specimens) GoogleMaps : MZUSP 122987 ( 28 specimens: juvenile female, 143 mm DW; juvenile female, 166 mm DW; juvenile female, 208 mm DW; juvenile female, 184 mm DW; juvenile female, 175 mm DW; juvenile female, 214 mm DW; juvenile male, 153 mm DW; embryo female, 154 mm DW; embryo male, 163 mm DW; female, 372 mm DW; juvenile male, 177 mm DW; juvenile male, 190 mm DW; juvenile male, 223 mm DW; juvenile male, 223 mm DW; juvenile male, 204 mm DW; juvenile female, 218 mm DW; juvenile male, 180 mm DW; adult male, not measured; juvenile male, 190 mm DW; male, 271 mm DW; juvenile male, 181 mm DW; juvenile male, 194 mm DW; male, 249 mm DW; male, 295 mm DW; male, 282 mm DW; male, 273 mm DW; juvenile female, 179 mm DW), Caiçara do Norte ; MZUSP 122988 (female, 317 mm DW), Caiçara do Norte ; MZUSP 122989 (female, 385 mm DW), Caiçara do Norte ; MZUSP 122990 (male, 296 mm DW), Caiçara do Norte ; MZUSP 122991 ( 2 specimens: juvenile female, 175 mm DW; juvenile male, 170 mm DW), Caiçara do Norte ; MZUSP 122992 (juvenile male, 174 mm DW), Caiçara do Norte ; MZUSP 122993 (female, 339 mm DW), Caiçara do Norte ; MZUSP 122994 (female, 486 mm DW), Caiçara do Norte ; MZUSP 122995 (female, 340 mm DW), Caiçara do Norte ; MZUSP 122996 (male, 316 mm DW), Caiçara do Norte ; MZUSP 122997 (female, 417 mm DW), Caiçara do Norte ; MZUSP 122998 (juvenile female, 186 mm DW), Caiçara do Norte ; MZUSP 122999 (male, 281 mm DW), Caiçara do Norte ; MZUSP 123000 (juvenile female, 252 mm DW), Caiçara do Norte ; RN FS179 (juvenile female, 267 mm DW, not preserved), Caiçara do Norte ; RN F133 (juvenile female, 163 mm DW, not preserved); RN F134 (juvenile female, 261 mm DW, not preserved); RN F135 (juvenile female, 285 mm DW, not preserved); RN F136 (juvenile female, 279 mm DW, not preserved); RN M102 (male, 310 mm DW, not preserved); RN M103 (juvenile male, 166 mm DW, not preserved); RN M104 (juvenile male, 217 mm DW, not preserved); RN M105 (male, 268 mm DW, not preserved); RN M106 (male, 285 mm DW, not preserved); RN M107 (juvenile male, 175 mm DW, not preserved). Paraíba, Brazil ( 14 specimens) : UERJ 1560 (not measured), Baía da Traição ; UERJ 1565 (not measured), Jacumã, Município do Conde ; UFPB 5/5/5 (2 specimens: juvenile female, 174 mm DW; juvenile male, 200 mm DW); UFPB 3040 (gravid female, 447 mm DW); UFPB 3041 (gravid female, 517 mm DW); UFPB 3488 (juvenile female, 182 mm DW); UFPB 5218 (adult male, 314 mm DW); UFPB 6077 (juvenile female, 275 mm DW); UFPB 6085 View Materials (juvenile male, 220 mm DW), Praia do Amor , Jacumã, Conde ; UFPB 6087 (juvenile female, 248 mm DW); UFPB 6525 (juvenile male, 182 mm DW); UFPB 6528 (juvenile male, 184 mm DW); UFPB 6674 View Materials (adult male, 281 mm DW), Praia da Penha, João Pessoa. Alagoas, Brazil ( 1 specimen) : MNHN A-8708 (dried, not measured), 10o00’00”S, 30o00’00”W. Sergipe, Brazil (1 specimen): UERJ 1523 (not measured), Praia do Pirambu . Bahia, Brazil ( 3 specimens): MNHN A-7972 (adult male, 355 mm DW), 12o58’01”S, 38o28’59”W GoogleMaps ; MNHN A-7973 (embryo, not measured), 12o58’01”S, 38o28’59”W; MZUSP 72920 View Materials (juvenile male, 198 mm DW), Ilhéus, Praia do Malhado e Malhadinho. Espírito Santo, Brazil ( 1 specimen) : MZUSP 9926 View Materials (adult male, 360 mm DW), in front of the Rio Doce River. Rio de Janeiro, Brazil ( 2 specimens) : ANSP 121582 (adult male, 346 mm DW), Atafona ; MZUSP 9735 (adult male, 353 mm DW), Atafona. Brazil, unspecified locality (1 specimens): ZMH 25815 (adult male, 326 mm DW).










No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Gymnura micrura ( Bloch & Schneider 1801 )
| Carvalho, Marcelo Rodrigues De 2017 |
Gymnura micrura
| McEachran 2002: 577 |
Gymnura micrura : Nishida, 1990 : 65
| Willems 2016: 36 |
| Fontenelle 2015: 7 |
| Garcia 2015: 4 |
| Ragno 2013: 57 |
| Yokota 2013: 1325 |
| Reis 2012: 217 |
| Yokota 2012: 1315 |
| Basilio 2009: 41 |
| Bornatowski 2009: 3 |
| Nunes 2009: 479 |
| Basilio 2008: 67 |
| Yokota 2007: 249 |
| Yokota 2006: 349 |
| Meneses 2005: 80 |
| Nunes 2005: 51 |
| Carvalho 2004: 10 |
| Furtado 2003: 8 |
| Menni 2000: 92 |
| Mazzoleni 1999: 144 |
| Lovejoy 1996: 214 |
| Nishida 1990: 65 |
Pteroplatea maclura : Puyo, 1936 : 79
| Puyo 1936: 79 |
Pteroplatea micrura
| Meek 1923: 87 |
Pteroplatea micrura : Metzelaar, 1919 : 8
| Metzelaar 1919: 8 |
Pteroplatea maclura
| Gunther 1870: 487 |
| Dumeril 1865: 614 |
| Muller 1841: 169 |
Pteroplatea micrura : Müller & Henle, 1841 : 169
| Pillay 1929: 353 |
| Annandale 1909: 39 |
| Alcock 1892: 1 |
| Wood-Mason 1891: 359 |
| Day 1889: 56 |
| Gunther 1870: 487 |
| Day 1865: 278 |
| Dumeril 1865: 613 |
| Blyth 1860: 29 |
| Cantor 1849: 1409 |
| Muller 1841: 169 |
Raja maclura : Lesueur, 1817 : 41
| Lesueur 1817: 41 |
Raja micrura
| Bloch 1801: 360 |