
Marmosops fuscatus ( Thomas, 1896 )
|
publication ID |
https://doi.org/ 10.1206/0003-0090-402.1.1 |
|
DOI |
https://doi.org/10.5281/zenodo.4630826 |
|
persistent identifier |
https://treatment.plazi.org/id/03A68972-9835-FFF6-06AD-71E6D639FB76 |
|
treatment provided by |
Felipe |
|
scientific name |
Marmosops fuscatus ( Thomas, 1896 ) |
| status |
|
Marmosops fuscatus ( Thomas, 1896) View in CoL
Figures 4B View FIG , 15 View FIG , 16 View FIG
Marmosa fuscata Thomas, 1896: 313 (original description).
Didelphys (Marmosa) fuscata: Trouessart, 1898: 1240 (name combination).
Marmosa (Marmosa) fuscata: Cabrera, 1919: 36 (name combination).
Marmosa cracens Handley and Gordon, 1979: 66 (original description). Type locality “near La Pastora, 14 km ENE Mirimire, Falcón, Venezuela.”
Marmosa crascens: Reig et al., 1985: 342 (incorrect subsequent spelling of M. cracens Handley and Gordon, 1979 ).
Marmosops cracens: Gardner and Creighton, 1989: 4 View in CoL (name combination).
Marmosops fuscatus: Gardner and Creighton, 1989: 4 View in CoL (first use of current binomial).
Marmosa (Marmosops) cracens: Pérez-Hernández et al., 1994: 35 (name combination).
Marmosa (Marmosops) fuscata: Pérez-Hernández et al., 1994: 37 (name combination); part (not including misidentified specimens of M. carri ).
Marmosops fuscatus fuscatus: Gardner and Creighton, 2008: 66 View in CoL (name combination); part (not including misidentified specimens of M. carri ).
TYPE MATERIAL: The holotype (by original designation) consists of the skin and skull of an old adult female ( BMNH 1896.11.1.6) collected by “S. Briceno” (= Salomón Briceño-Gabaldón, a commercial collector) at 1630 m elevation along the Río Albarregas (misspelled “Abbaregas” by Thomas, 1896) near the city of Mérida (fig. 17: locality 114) in the Venezuelan state of Mérida on 6 April 1896.
DISTRIBUTION, HABITATS, AND SYMPATRY: Examined specimens of Marmosops fuscatus are from four Venezuelan localities, two in the Mérida Andes (Cordillera de Mérida), one in the Serranía de San Luis of western Falcón, and one in the Caribbean coastal lowlands of eastern Fal-
cón (fig. 17). Although recorded elevations range from 125 to 2350 m above sea level, capture habitats seem to be either premontane or montane rain forest. 6 Previous reports of this species from Colombia (e.g., Gardner and Creighton, 2008: map 28) were based on misidentified material (see Remarks, below). Marmosops fuscatus is not known to occur sympatrically with any congeneric species, although it could be expected to do so with M. carri and M. ojastii , which have adjacent or overlapping geographic ranges.
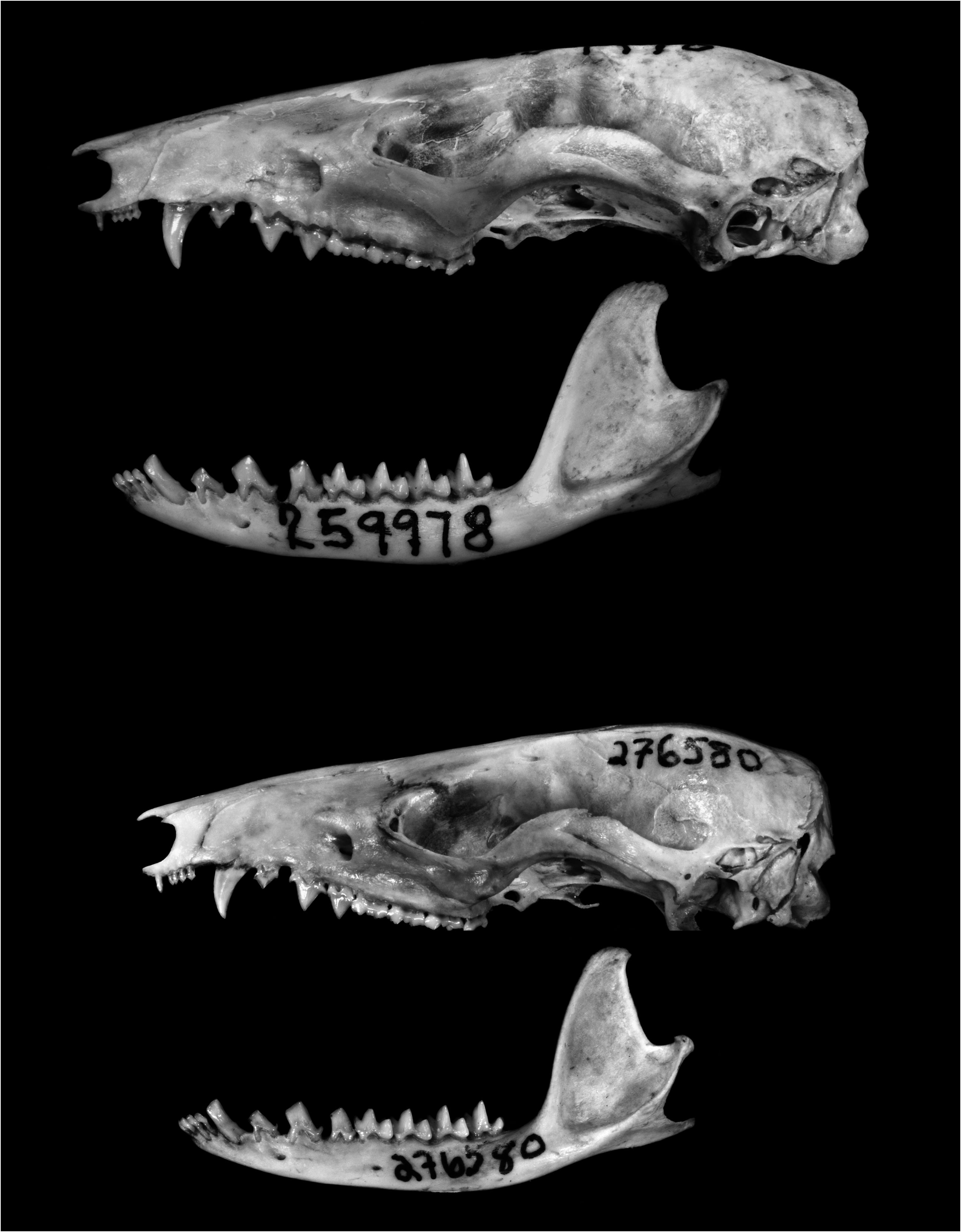
DESCRIPTION: Body pelage dark grayish brown (near Sepia) middorsally, indistinctly paler laterally, and about 7–9 mm long at midback (shorter in specimens from lower elevations than in those from the Mérida Andes); ventral pelage superficially whitish but uniformly gray based except for the apex of the chin, oral margins, and scrotum (which are covered with selfwhitish hairs). (One examined specimen, AMNH 276509, has small patches of self-white hairs along the inner thighs.) Manus covered dorsally with uniformly pale hairs (the metacarpals not contrasting in color with the digits); lateral carpal tubercles bladelike in all examined adult males. Mammary formula unknown. Tail longer than combined length of head and body (mean LT/HBL × 100 = 110%); dorsal caudal surface dark, but indistinctly paler distally in some specimens; ventral surface distinctly paler.
Nasal bones long (extending well behind the lacrimals) and uniformly narrow (without distinct lateral expansion at the maxillary-frontal suture). Interorbital region broad, with rounded supraorbital margins in females and young adult males, but relatively narrower and tending to develop squared supraorbital margins in large adult males; postorbital processes absent or 6 Handley and Gordon (1979) reported that the type series of Marmosa cracens was collected between 125 and 170 m above sea level on “the steep, moist, north slope of an isolated, low mountain” near the Caribbean coast. Vegetation of distinctly montane character can extend downslope to within 100 m above sea level on such isolated peaks, which intercept moisture-laden sea breezes and are often shrouded in low-lying clouds. The cloud forest (montane rain forest) vegetation at capture localities in the Serranía de San Luis was briefly described by Anderson et al. (2012).
indistinct. Lacrimal foramina variable (the upper foramen is usually not visible in lateral view, but the lower foramen can be either laterally visible or concealed inside the anterior orbital margin); 7 zygomatic process of squamosal broadly overlapped dorsally by the jugal. Palatine fenestrae present or absent (when present these can appear as a single small perforation on each side or as several small irregular perforations on both sides). Dorsolateral margin of ethmoid foramen usually formed by the frontal.
Upper canine (C1) sexually dimorphic (longer and unicuspid in males, shorter and with posterior accessory cusp in young female specimens with unworn teeth). Upper third molar (M3) anterolabial cingulum discontinuous with preprotocrista (anterior cingulum incomplete). Lower canine (c1) sexually dimorphic (erect, without a posterior accessory cusp, and taller than p 1 in males versus procumbent, with a posterior accessory cusp, and subequal in height to p 1 in females); c1 anterolingual accessory cusp usually absent in both sexes. Entoconid of m1 subequal in height to adjacent m2 paraconid; unworn m4 talonid with three distinct cusps.
COMPARISONS: Comparisons between Marmosops fuscatus and M. carri have already been described (see above).
Marmosops fuscatus is substantially larger than M. handleyi in head-and-body length and has a proportionately shorter tail (tables 5, 6). Externally the two species differ in the same characters that distinguish M. carri from M. handleyi , namely dorsal pelage coloration (dark grayish brown in fuscatus , dark brown in handleyi ) and forefoot coloration (the manus is entirely whitish in fuscatus , whereas the pale digits contrast with the dark metacarpals in handleyi ). Marmosops fuscatus also averages larger than M. handleyi in most craniodental dimen-
7 The lower lacrimal foramen is laterally visible in some specimens (e.g., BMNH 3.1.5.2, 3.1.5.3, FMNH 22174, USNM 442719) but not in others (e.g., AMNH 276580, 276509). The specimens in which this foramen is visible are the largest in overall size and have the most heavily ossified skulls, but we lack sufficient material to determine whether or not this trait is age correlated.
sions, including four measurements (CBL, ZB, PL, MTR, and WM3) that exhibit nonoverlapping variation in same-sex comparisons. These species also differ in some of the same qualitative craniodental characters that distinguish M. carri and M. handleyi ; in particular, M. fuscatus has much narrower nasals than M. handleyi (in which these bones are conspicuously expanded laterally at the maxillary-frontal suture), and males of M. fuscatus have unicuspid C1 and c1, whereas these teeth consistently have a small posterior accessory cusp in males of M. handleyi . Additionally, the dorsolateral margin of the ethmoid foramen is usually formed by the frontal in M. fuscatus , whereas the dorsolateral margin of this foramen is formed by the orbitosphenoid in M. handleyi . Lastly, the distinct lingual accessory cusp of c1 that appears to be consistently present in M. handleyi is absent in M. fuscatus .
Marmosops fuscatus is substantially larger than M. invictus in all external dimensions. In qualitative external characters, M. fuscatus is most readily distinguished from M. invictus by its paler dorsal coloration (the dorsal pelage of invictus is consistently much darker), by its entirely pale forefeet (the metacarpals are dark in M. invictus ), and by its more distinctly bicolored tail (the tail is only indistinctly paler ventrally than dorsally in M. invictus ). Marmosops fuscatus averages larger than M. invictus in most craniodental dimensions, including five measurements (CBL, NL, ZB, PL, MTR) that exhibit nonoverlapping variation in same-sex comparisons. Craniodental qualitative differences between these quite dissimilar species include the shape of the nasals (uniformly narrow in M. fuscatus , laterally expanded in M. invictus ), dorsolateral margin of the ethmoid foramen (usually formed by the frontal in M. fuscatus , by the orbitosphenoid in M. invictus ), C1 posterior accessory cusp (absent in males of M. fuscatus , present in both sexes of M. invictus ), M3 anterior cingulum (incomplete in M. fuscatus , narrowly complete in M. invictus ), c1 morphology (erect and unicuspid in male M. fuscatus , procumbent and premolariform in M. invictus ), c1 lingual accessory cusp (absent in M. fuscatus , usually present in M. invictus ), and m1 entoconid (subequal to adjacent m2 paraconid in fuscatus , shorter than m2 paraconid in M. invictus ).
REMARKS: Marmosa perfusca , a nominal species that Thomas (1924) described from the Cordillera Oriental of Colombia, was synonymized with M. fuscatus by Tate (1933: 173), and some authors (e.g., Gardner and Creighton, 2008: 66) have treated it as a valid subspecies (“ Marmosops fuscatus perfuscus ”). However, the type of perfusca (BMNH 1923.11.13.18, consisting of the skin and skull of an old adult female) has posteriorly widened nasals, a short subsquamosal foramen (not exposing the petrosal behind the sulcus of the prootic sinus), and only one antebrachial vibrissa. In these and other characters it closely resembles Marmosops caucae ( Thomas, 1900) , a member of the nominotypical subgenus of Marmosops . A second Colombian specimen that Tate (1933) referred to Marmosa fuscata is AMNH 34641, also from the Cordillera Oriental; 8 like BMNH 1923.11.13.18, this specimen appears to represent M. caucae . To the best of our knowledge, there are no valid records of Marmosops fuscatus from Colombia.
Handley and Gordon (1979: 67) described Marmosa cracens as “in almost every respect of pelage and skull character ... a miniature replica of M. fuscata , distinguishable from it only by its small size and the short, broad shape of its palate.” Indeed, the male holotype of cracens (USNM 418503) is substantially smaller than any adult male specimen of Marmosops fuscatus that we measured. For example, the condylobasal length of the skull of USNM 418503 is 28.4 mm, whereas the range of condylobasal lengths of five adult male M. fuscatus is 31.8–35.4 mm. How-
8 This specimen was collected by G. O’Connell on 4 April 1913 at “El Roble,” a locality that Tate (1933: 173) erroneously placed in Departamento Cauca and that Gardner and Creighton (2008: 66) located in Departamento Quindío (on the western slope of the Cordillera Central; Paynter, 1997). However, Geoffroy O’Connell was a member of the 1913 AMNH expedition that worked at a different place called El Roble, “a posada on the trail from Bogotá to Fusugasugá [= Fusagasugá]” on the western slope of the Cordillera Oriental ( Chapman, 1917: 645); according to Paynter (1997), this locality is near 4°23′N, 74°19′W.
ever, this size difference is plausibly explained by ontogeny: the type series of cracens consists of subadults (with incompletely erupted P3s), not young adults (contra Handley and Gordon, 1979). The subadult holotype and paratype of cracens do have relatively wider palates (PB/PL = 0.57–0.59) than topotypical adult fuscatus (0.51– 0.53), but relative width of the palate is another ontogenetically variable trait because didelphid palates become proportionately longer and narrower with advancing age ( Abdala et al., 2001). Lastly, we ( Díaz-Nieto et al., 2016a) amplified partial (579–619 bp) cytochrome- b sequences from the holotype and a paratype of cracens , which differ by only 2.2% (uncorrected) from a partial (1008 bp) cytochrome- b sequence that we amplified from a topotype of M. fuscatus (BMNH 1903.1.5.2). Because this genetic distance is well within the range of values commonly obtained from comparisons of conspecific sequences of other mammals, and because the phenotypic differences that Handley and Gordon (1979) alleged between cracens and fuscatus seem to be attributable to the immaturity of their type series, we treat the former as a subjective junior synonym.
SPECIMENS EXAMINED (N = 8): Venezuela — Falcón, Near La Pastora ( USNM 418503 About USNM [holotype of cracens ], 442719), Serranía de San Luís ( AMNH 276509 About AMNH , 276580 About AMNH ) ; Mérida, La Azulita ( FMNH 22174 About FMNH ), Mérida ( BMNH 1896.11 .1.6 [holotype], 1903.1.5.2, 1903.1.5.3) .


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Marmosops fuscatus ( Thomas, 1896 )
| Díaz-Nieto, Juan F. & Voss, Robert S. 2016 |
Marmosops fuscatus fuscatus: Gardner and Creighton, 2008: 66
| Gardner, A. L. & G. K. Creighton 2008: 66 |
Marmosa (Marmosops) cracens: Pérez-Hernández et al., 1994: 35
| Perez-Hernandez, R. & P. Soriano & D. Lew 1994: 35 |
Marmosa (Marmosops) fuscata: Pérez-Hernández et al., 1994: 37
| Perez-Hernandez, R. & P. Soriano & D. Lew 1994: 37 |
Marmosops cracens:
| Gardner, A. L. & G. K. Creighton 1989: 4 |
Marmosops fuscatus: Gardner and Creighton, 1989: 4
| Gardner, A. L. & G. K. Creighton 1989: 4 |
Marmosa crascens:
| Reig, O. A. & J. A. W. Kirsch & L. G. Marshall 1985: 342 |
Marmosa cracens
| Handley, C. O., Jr. & L. K Gordon 1979: 66 |
Marmosa (Marmosa) fuscata:
| Cabrera, A. 1919: 36 |
Didelphys (Marmosa) fuscata:
| Trouessart, E. - L. 1898: 1240 |
Marmosa fuscata
| Thomas, O. 1896: 313 |