
Hyalonema (Cyliconemaoida) alucia, Moser & Moraes & Castello-Branco & Pequeno & Muricy, 2022
|
publication ID |
https://doi.org/ 10.11646/zootaxa.5105.1.4 |
|
publication LSID |
lsid:zoobank.org:pub:B5C60031-4303-4CBA-A59C-22AE34B73631 |
|
DOI |
https://doi.org/10.5281/zenodo.6335976 |
|
persistent identifier |
https://treatment.plazi.org/id/03A687C1-FFEE-137F-B1B1-FBE549AA2FCF |
|
treatment provided by |
Plazi |
|
scientific name |
Hyalonema (Cyliconemaoida) alucia |
| status |
sp. nov. |
Hyalonema (Cyliconemaoida) alucia View in CoL sp. nov.
( Figures 2–5 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 )
Material examined. Holotype MNRJ 21298 View Materials , Brazil, off Pará State, upper slope, 01°31.104`N – 046°43.929`W, 378 m depth, coll. F Moraes / MV Alucia, submersible Deep Rover, 18.vii.2017. GoogleMaps
Diagnosis. Hyalonema (Cyliconemaoida) with caliciform body, with a disorganized sieve plate in the apical surface. Choanosomal spicules are mainly smooth diactins. Dermalia are pinular pentactins with relatively short, straight pinular ray directed outwards. Atrialia are pinular pentactins, smooth pentactins and hexactins, and spined microxypentactins and microxyhexactins. Amphidiscs in three size categories similar in shape, all with umbels longer than broad and smooth teeth largely parallel to the shaft. Basalia diactins apparently without anchors in the ends.
Etymology. The specific epithet alucia is a noun in apposition given in honor of the MV Alucia and its crew, responsible for the collection of this species.
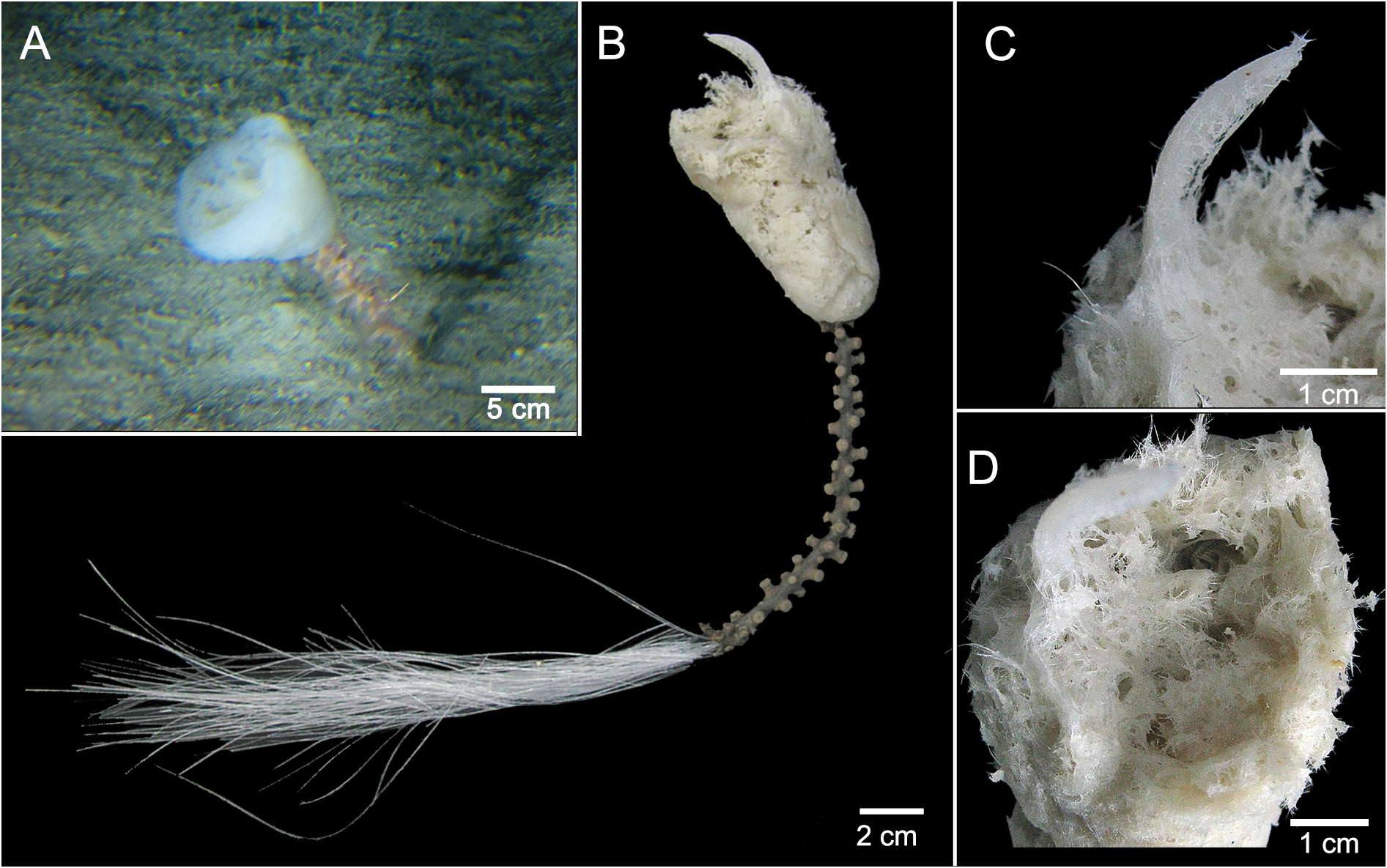
Description. Sponge lophophytous, caliciform, measuring 45.5 cm in total length; body 8.7 cm long by 5 cm wide and basal tuft 37.5 cm long ( Fig. 2A, B View FIGURE 2 ). Apical surface with a central cone, small in vivo but displaced to one side after fixation ( Fig. 2A–C View FIGURE 2 ) and a disorganized sieve-plate, damaged after collection, with abundant circular openings 2–8 mm in diameter ( Fig. 2D View FIGURE 2 ). External surface even, microhispid. Color white in vivo, becoming white to cream in ethanol. Consistency of the body soft, easy to tear. Basal tuft resistant, fully covered with zoanthids on its exposed portion, from the base of the body down to 12.5 cm of the tuft ( Fig. 2A, B View FIGURE 2 ).
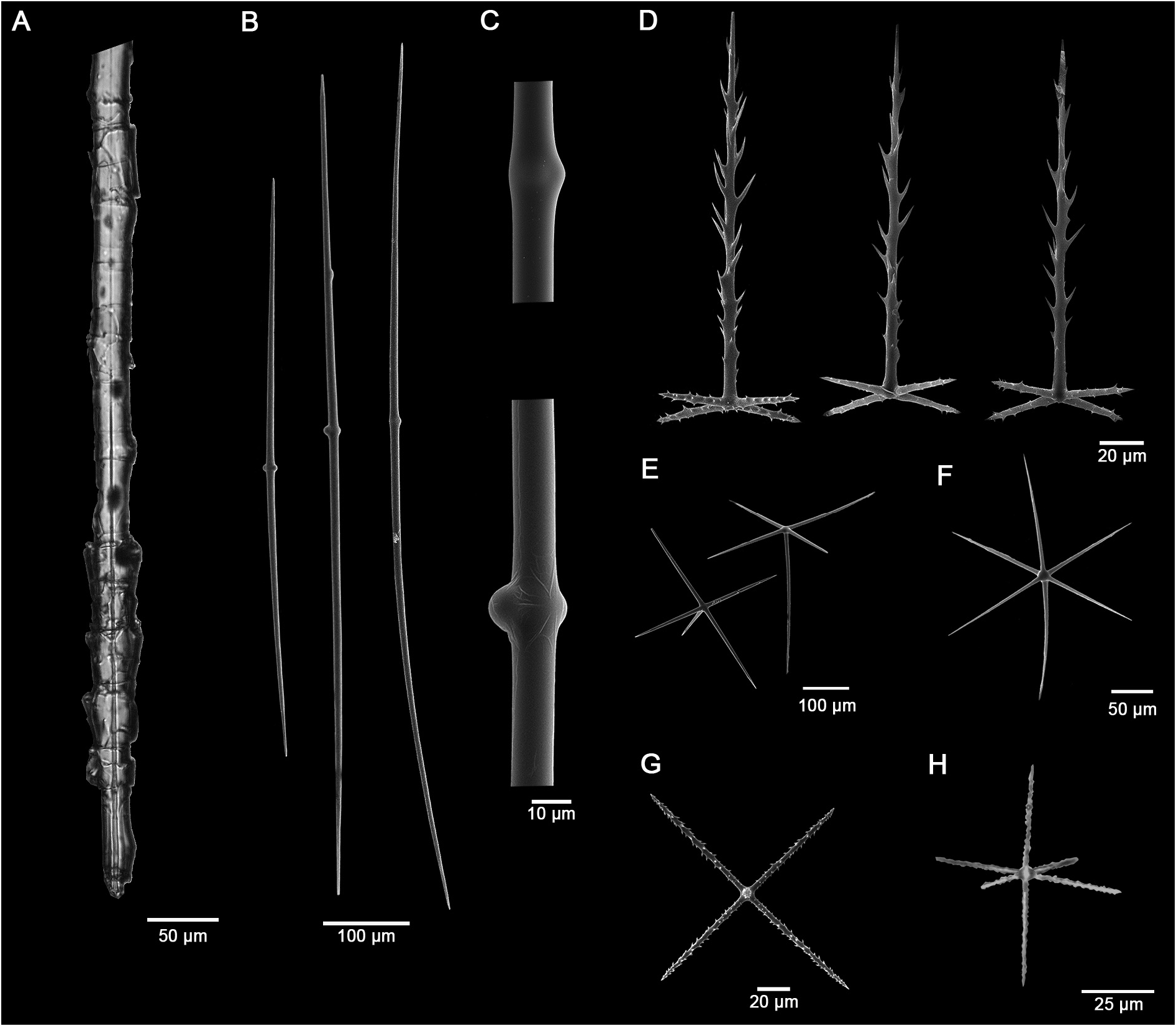
Skeleton: Choanosome mainly composed of abundant, disorganized diactins ( Fig. 3A View FIGURE 3 ). Dermalia includes diactins tangential to the surface or in tufts and pinular pentactins with the pinular rays directed outwards ( Fig. 3B View FIGURE 3 ). Atrialia contain diactins, pinular pentactins, spined hexactins, and smooth pentactins and hexactins. Canalaria composed of smooth pentactins and hexactins, slightly smaller than in atrialia, and abundant pinular pentactins with the pinular ray directed to the interior of the canals. Basalia diactins apparently terminating in sharp ends, without visible anchors or teeth even when not broken. Acanthophores, pinular diactins and ambuncinates absent.
Spicules: Megascleres are basal diactins, smooth choanosomal diactins, dermal and atrial pinular pentactins, atrial smooth pentactins and hexactins. Microscleres are atrial spined microoxypentactins and microoxyhexactins plus macramphidiscs, mesamphidiscs and micramphidiscs ( Figs 4 View FIGURE 4 , 5 View FIGURE 5 ; Table 1 View TABLE 1 ).
Diactins of the basalia with irregular striations formed by terminations of boxed concentric layers of silica. Ends acerate, not anchored. Size up to 390 mm long by 20–200 µm wide ( Fig. 4A View FIGURE 4 ).
Choanosomal diactins smooth, straight to lightly curved, widening in the central region with sharp or rounded ends; 560–952–1500 µm in length by 7.5–13.8–22.5 µm in width ( Fig. 4 View FIGURE 4 B-C).
Both dermal and atrial pinular pentactins with straight, relatively short, whip-like pinular ray with relatively long spines, shaft thickest at the base, 75.0–157.0–230.5 µm in length by 2.5–3.7–5.0 µm in width ( Fig. 4D View FIGURE 4 ). Tangential rays shorter than pinular ray, microspined; diameter of the basis 50.0–69.5–100.0 µm.
Atrial pentactins symmetrical, with smooth, straight rays. Proximal ray 100.0–325.5–460.0 µm in length, tangential rays 125.2–233.2–445.0 µm long by 5.0–8.8–15.0 µm in width ( Fig. 4E View FIGURE 4 ).
Atrial hexactins smooth ( Fig. 4F View FIGURE 4 ) with usually straight and occasionally slightly curved rays. Rays symmetrical, with center sometimes slightly enlarged. Proximal ray 140.0–276.5–420.0 µm length, distal ray 100.0–235.5–490.0 µm in length, tangential rays 100.0–222.8–425.0 µm in length and 4.5–7.9–17.5 µm in width.
Microxypentactins rare, microspined, with symmetric tangential rays longer than the proximal ray. Proximal ray length 30.0–50.8–90.0 µm, tangential rays 63.8–75.9–92.5 µm in length and 2.5–2.7–4.5 µm in width ( Fig. 4G View FIGURE 4 ).
Microxyhexactins microspined, asymmetrical, with proximal and distal rays longer than the tangential ones ( Fig. 4H View FIGURE 4 ). Proximal ray 37.5–67.5–100.0 µm in length, distal ray 35.0–64.0–82.5 µm in length, tangential rays 45.0–64.8–98.8 µm length by 2.5–2.7–3.3 µm in width.
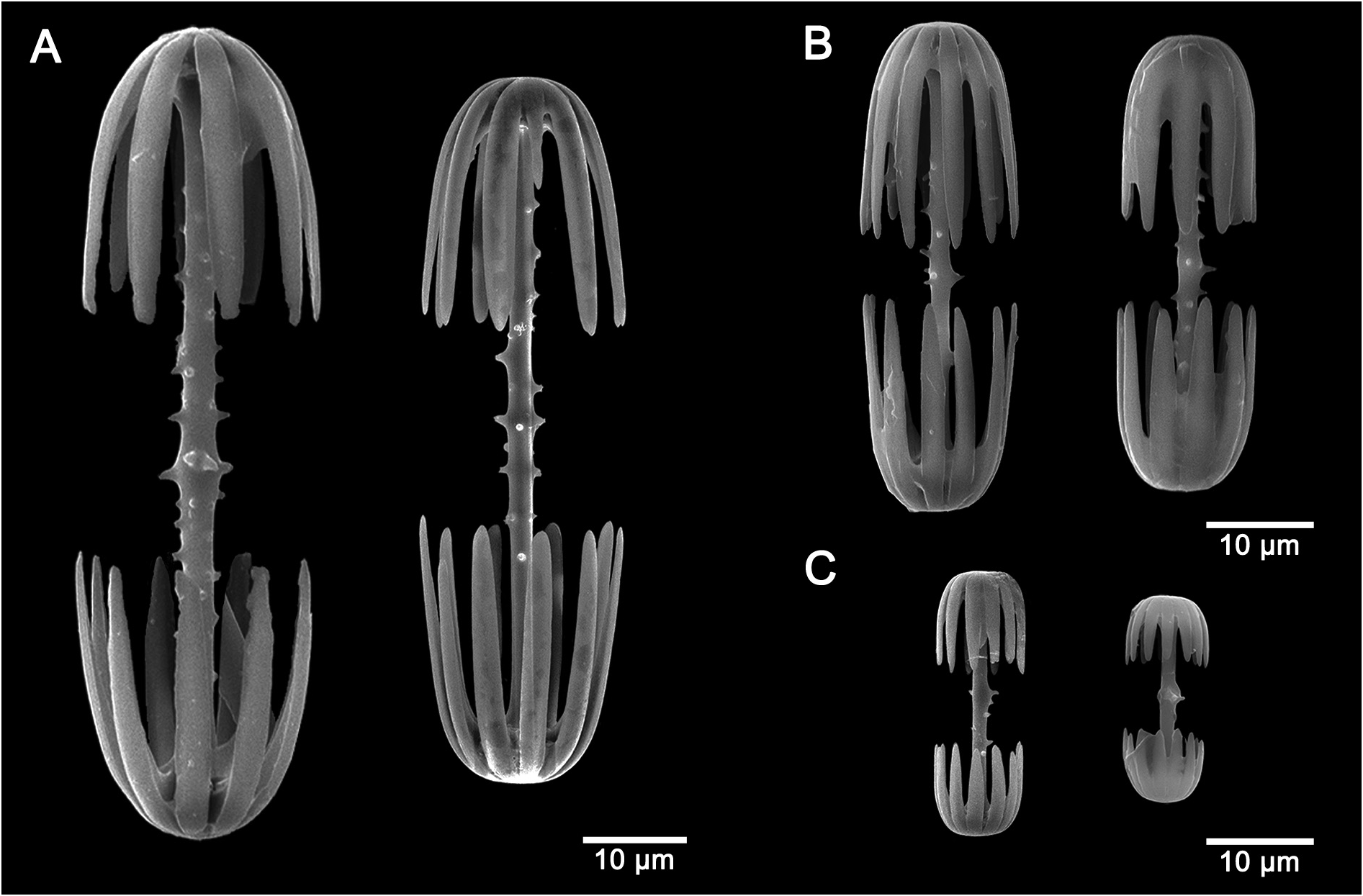
Macramphidiscs ( Fig. 5A View FIGURE 5 ) with smooth umbels longer than broad or as long as broad; umbel length 50.0–64.0– 80.0 µm and umbel width 23.0–54.0–88.0 µm. Shaft spiny to tuberculated, 82.5–152.5–250.0 µm in total length and by 2.5–7.0–17.5 µm in width.
Mesamphidiscs ( Fig. 5B View FIGURE 5 ) with smooth umbels covering almost the whole spicule length; umbel length 17.5– 23.6–30.0 µm, umbel width 12.5–19.0–22.5 µm. Shaft spiny to tuberculated, 40.0–56.3–70.0 µm in total length by 2.0–2.4–2.5 µm in width.
Micramphidiscs ( Fig. 5C View FIGURE 5 ) with smooth umbels, broader than long; umbel length 5.0–8.2–15.0 µm, umbel width 7.5–8.0–10.0 µm. Shaft spiny to tuberculated, 15.0–21.0–32.5 µm in total length by 1.0 µm in width.
Geographical and Bathymetric Distribution: Only known from the upper slope off Pará State, Northern Brazil, 378 m depth .
Ecology. The single specimen was anchored by the basal spicule tuft on muddy sand bottom ( Fig. 2A View FIGURE 2 ). Zoanthids grow over all parts of the basal spicule tuft outside the sediment ( Fig. 2A, B View FIGURE 2 ).
Taxonomic remarks. The genus Hyalonema is divided in 12 subgenera ( Tabachnick & Menshenina 2002; de Voogd et al. 2021). The new species is allocated in the subgenus Cyliconemaoida (formerly Leptonema Lendenfeld, 1915 ) due to the presence of amphidiscs with umbels longer than broad and pinular pentactins with shaft thickest at base and whip–like shape ( Tabachnick & Menshenina 2002; Dohrmann 2017).
Only three species of Hyalonema were previously reported from Brazil, none from subgenus H. ( Cyliconemaoida ): Hyalonema (Coscinonema) schmidti Schulze, 1899 ; H. (Cyliconema) conqueror Tabachnick et al., 2009 and H. (Prionema) dufresnei Tabachnick et al., 2009 . Of these, Hyalonema (Cyliconema) conqueror seems closer to the new species in body shape and spiculation, but it differs by having acanthophores stauractins and pinular diactins, which are absent in Hyalonema (Cyliconemaoida) alucia sp. nov. In addition, the amphidiscs of the new species present umbels longer than broad, while in H. (Cyliconema) they are broader than long ( Tabachnick & Menshenina 2002).
The subgenus Hyalonema (Cyliconemaoida) currently contains 10 accepted species (Dohrmann 2017; de Voogd et al. 2021), which differ from the new species by shape and spiculation. Hyalonema (C.) choaniferum ( Lévi, 1964) , H. (C.) ovuliferum Schulze, 1899 sensu Kersken et al. (2018) and H. (C.) urna Schulze, 1904 differ by having a spherical to ovoid shape, with an apical oscular tube surrounded by marginalia. In addition, H. (C.) ovuliferum has ovoid to ellipsoidal macramphidiscs and pinular diactins; H. (C.) urna has acanthophores and marginalia diactins; and H. (C.) choaniferum has ovoid macramphidics with long-teethed umbels and pinular pentactins with long proximal ray. Hyalonema (C.) campanula Lendenfeld, 1915; H. (C.) ovuliferum and H. (C.) divergens Schulze, 1887 have pinular diactins, absent in the new species.
The new species is characterized by the presence of spined microxypentactins, the proximal ray of the pinular pentactins being straight and with relatively long spines, and by the almost rectangular shape of its macramphidiscs, in which the umbel teeth become parallel to the shaft. In most other species of Hyalonema (Cyliconemaoida) the macramphidiscs umbels are ovoid [ H. (C.) ovuliferum and H. (C.) choaniferum ( Lévi, 1964) ], campanulate, with straight teeth diverging outwards [H. (C.) campanula and H. (C.) divergens ], or hemispherical to bell-shaped [H. (C.) acuferum Schulze, 1893; H. (C.) solutum Schulze, 1904; H. (C.) flagelliferum Ijima, 1927; H. (C.) spatha Tabachnick & Lévi, 2000 ; H. (C.) urna]. The only other species with almost rectangular macramphidiscs in the subgenus is H. (C.) lusitanicum Barboza du Bocage, 1864 , which however differs by having also stauractins and parenchymal monactins.
Hyalonema (C.) flagelliferum from the Banda Sea differs from the new species by the much longer proximal ray of the pinular pentactins (500–800 µm long), the presence of acanthophores (stauractins), the absence of microxypentactins and microxyhexactins, and by the larger size and different shape of its macramphidiscs (220–418 µm long, with bell-shaped umbels; Ijima 1927).
Hyalonema (C.) spatha from New Caledonia differs by the abundant canalar depressions on the apical surface, the presence of acanthophores (sword-like stauractins) and pinular diactins, the absence of hexactins, microxypentactins, microxyhexactins and mesamphidiscs, and by the smaller size and different shape of its macramphidiscs (25–148 µm long, with divergent umbel teeth; Tabachnick & Lévi 2000).
Hyalonema (Cyliconemaoida) alucia sp. nov. is the first species of the subgenus described in the Western Atlantic, while all others occur in the Indian and Pacific oceans. It is also the shallowest record of the subgenus, at 378 m depth, while H. (C.) spatha occurs from 489 to 1005 m depth ( Tabachnick & Lévi 2000) and the other species occur from 1527 to 5300 m depth ( Schulze 1887, 1893, 1899, 1904; Lendenfeld 1915; Ijima 1927; Lévi 1964; Boury-Esnault et al. 1994; Kersken et al. 2018).

| MV |
University of Montana Museum |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
|
|
SubGenus |
Hyalonema |