
Pseudomesochra tatianae Drzycimski, 1968
|
publication ID |
https://doi.org/ 10.11646/zootaxa.5051.1.16 |
|
publication LSID |
lsid:zoobank.org:pub:BDE521C5-BB26-4445-8826-9CA5B2A00A03 |
|
DOI |
https://doi.org/10.5281/zenodo.5576300 |
|
persistent identifier |
https://treatment.plazi.org/id/03A587F5-FFA0-1A7D-FF4F-F9688115FD52 |
|
treatment provided by |
Plazi |
|
scientific name |
Pseudomesochra tatianae Drzycimski, 1968 |
| status |
|
Pseudomesochra tatianae Drzycimski, 1968
( Figs. 8–11 View FIGURE 8 View FIGURE 9 View FIGURE 10 View FIGURE 11 )
Type locality. Norway, Norway, Korsfjord , 60º12’24”N, 05º13’06”E; mud at 512 m depth GoogleMaps .
Original description. Drzycimski (1968): 18–20, Abb. 2.
Material examined. 1 ♀ (reg. no. NIBRIV0000853917 ) dissected on seven slides , 1 ♀ dissected on six slides (reg. no. NIBRIV0000853918 ), and 2 ♀♀ (reg. no. MInRB-Hr48-L001) preserved together in ethanol. Collected in Southern Sea of Korea, 33º30’N, 124º00’E (St. 1 in Fig. 1 View FIGURE 1 ), 70.8 m depth, 05 June 2015 GoogleMaps .
1902: Plate XXIV, Fig. 35).
3: Lang’s (1948: 641) statement on the armature of P2 is mistakenly based on Sars’ (1906: plate CXXV) illustration of P3; neither P2 nor P4 were figured by Sars but his text description (p. 193) revealed that P2–P4 exp-1 and -3 lack an inner seta and the armature pattern of the endopods is 1.221. Willen’s (1996: 103) statement that P. gertwilleni shares with P. divaricata the presence of an inner seta on P2–P4 exp-3 is therefore wrong. Coull (1973: Table II) and Vasconcelos et al. (2008) assumed that the exopodal pattern for P2– P4 was 0.1.023 but this requires confirmation. Unlike other known congeners both members of the female P5 are medially fused in P. divaricata (see Sars 1906: plate CXXV).
4: As pointed out by Lang (1948) Brady’s (1910) illustrations of P2–P3 are probably incorrect and the armature patterns therefore potentially misleading.
5: Sars (1911: Supplement Plate 25) shows a P2 with three inner setae (the distal one being smaller than the other two) on exp-3 resulting in a 323 formula. According to Huys & Boxshall (1991: 119) the maximum number of elements on this segment is seven, the presence of only two inner setae being an apomorphy for the Harpacticoida . It is therefore concluded that Sars must have figured P4 instead of P2; this mistake was adopted by Lang (1948), Coull (1973) and Vasconcelos et al. (2008).
6: Lang (1936b: 174) considered this species a junior subjective synonym of P. longifurcata . Characters of both descriptions ( Scott 1902; Sars 1920), such as the ornamentation of the caudal ramus, morphology of ♀ P5 and possibly armature of P2–P3, were combined in Lang’s (1948) diagnosis of P. longifurcata and used unwisely in subsequent identification keys ( Coull 1973; Willen 1996) (see also 2). This course of action also resulted in the unjustified acceptance of a variable armature of P4 enp-2 ( Coull 1973: Table II). Pseudomesochra affinis exhibits only one (vs two in P. longifurcata ) inner seta(e) on P4 enp-2 and displays (vs lacks) a proximal spinular patch along the inner margin of the caudal ramus ( Sars 1920: Plate XXXIV). Additional differences are found in the number of mandibular basal setae, the ornamentation and armature of the maxilliped, and in the shape of the caudal ramus (straight vs concave outer margin). Based on these differences P. affinis is removed from its synonymy with P. longifurcata and formally reinstated as a valid species.
7: According to Lang (1936b: 176, Abb. 7) P3–P4 enp-3 have only one apical seta, resulting in a 211 armature formula. In his monograph however this formula was changed to 221 ( Lang 1948: 593, Table X); it is unclear whether this is a typographical error or the result of new observations. Note that Coull (1973: Table II, 603) and Wells (2007: 689) used the 221 pattern in their keys and so did Vasconcelos et al. (2008) in their Table 1 View TABLE 1 despite Drzycimski (1968: 20) having pointed out the discrepancy.
8: Becker & Schriever (1979: 52) erroneously listed the armature formula of P2 exopod as 1.222.
9: Becker & Schriever’s (1979: 52) description of P3 shows a discrepancy between the text (1.1.221) and the illustration (1.1.211); the latter is adopted here.
10: Willen (1996) describes and illustrates the ♀ antennule as 6-segmented but erroneously uses the 7-segmented condition in her identification key (couplet 10).
11: According to Willen & Dittmar’s (2009) diagnosis of Keraia the antennule is 5-segmented; however, it was denoted as 6-segmented by Smirnov (1946) in his text description of K. tamara (although only the first four segments were illustrated in full). Smirnov’s (1946) armature formulae of the endopods of P2 (1.220) and P3–P4 (1.320) were reinterpreted by Coull (1973) as 1.121 and 1.221, respectively. Smirnov also lists the armature pattern of P4 exopod as 1.1.323 but only illustrates it as 1.1.223. The former is adopted here since it corresponds to the one found in Coull’s (1973) specimens from the deep sea off North Carolina. One male was reported by Smirnov (1946) —the only one in the genus Keraia so far—but no morphological information was given.
12: These species—both described from the Southern Atlantic—are extremely similar and may well be conspecific. They are here provisionally maintained as separate species based on small differences in cephalothoracic length, length ratio of apical setae on the antennary exopod, relative length of P1 endopodal segments and setal arrangement on the ♀ P5 endopodal lobe.
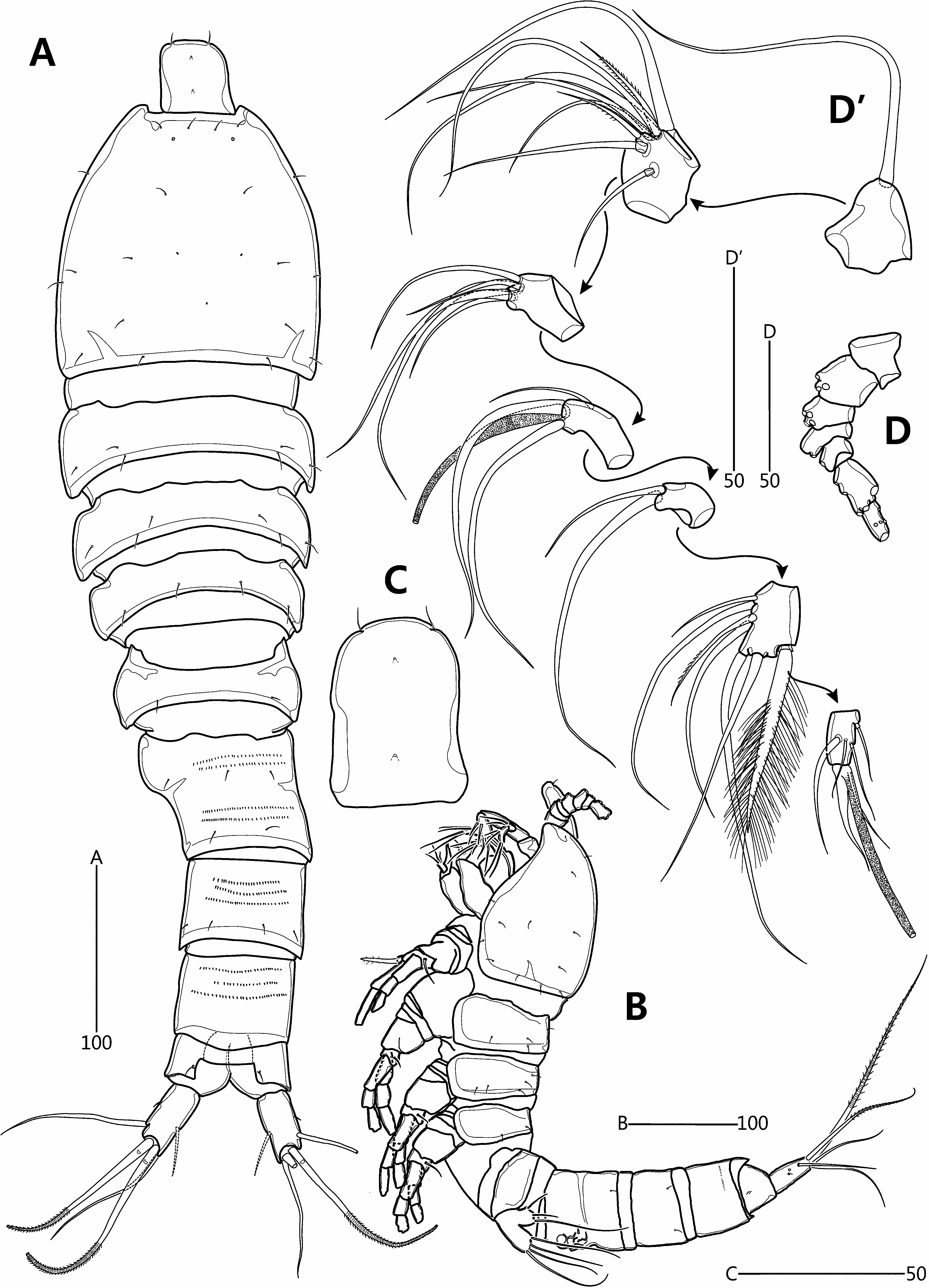
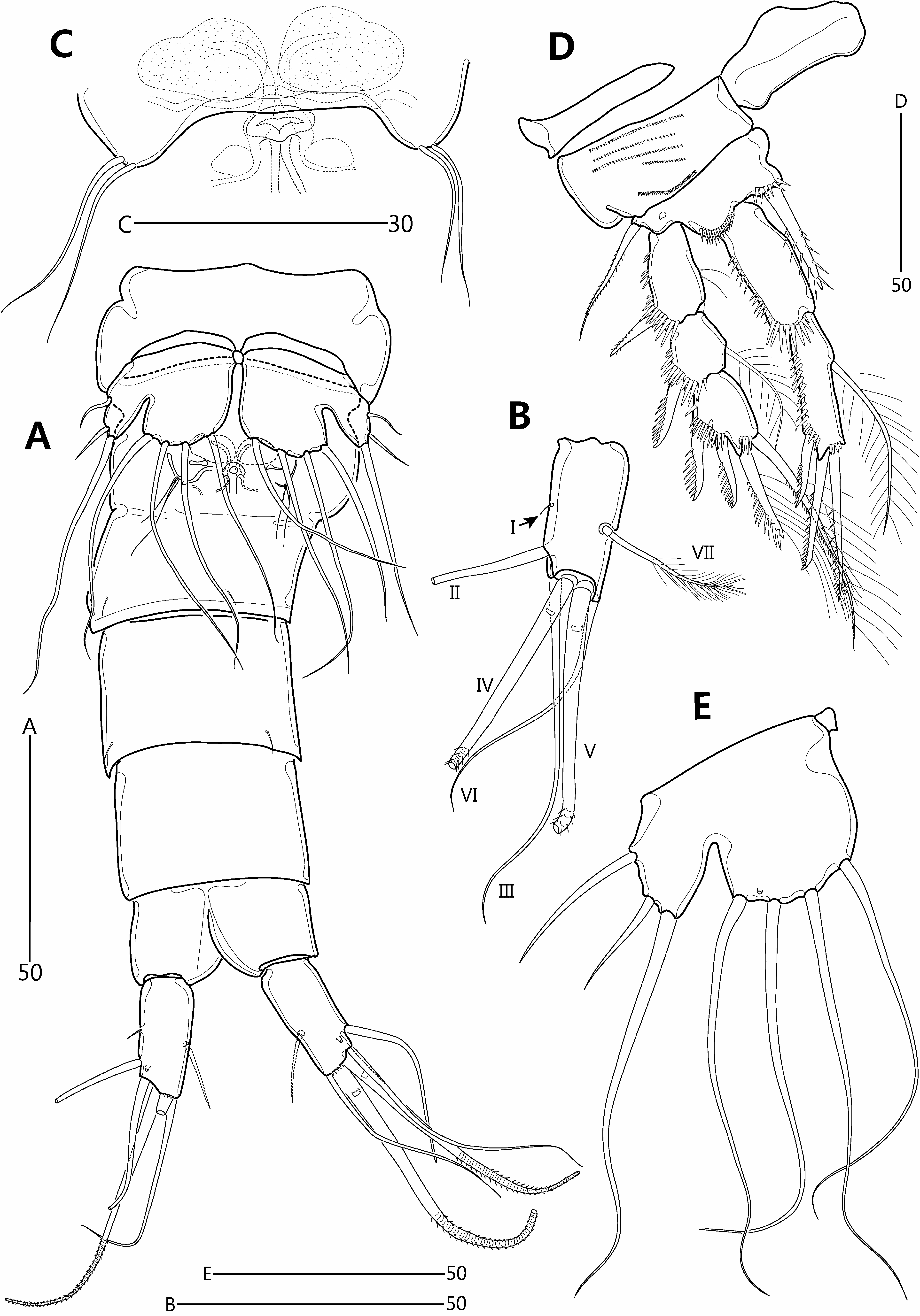
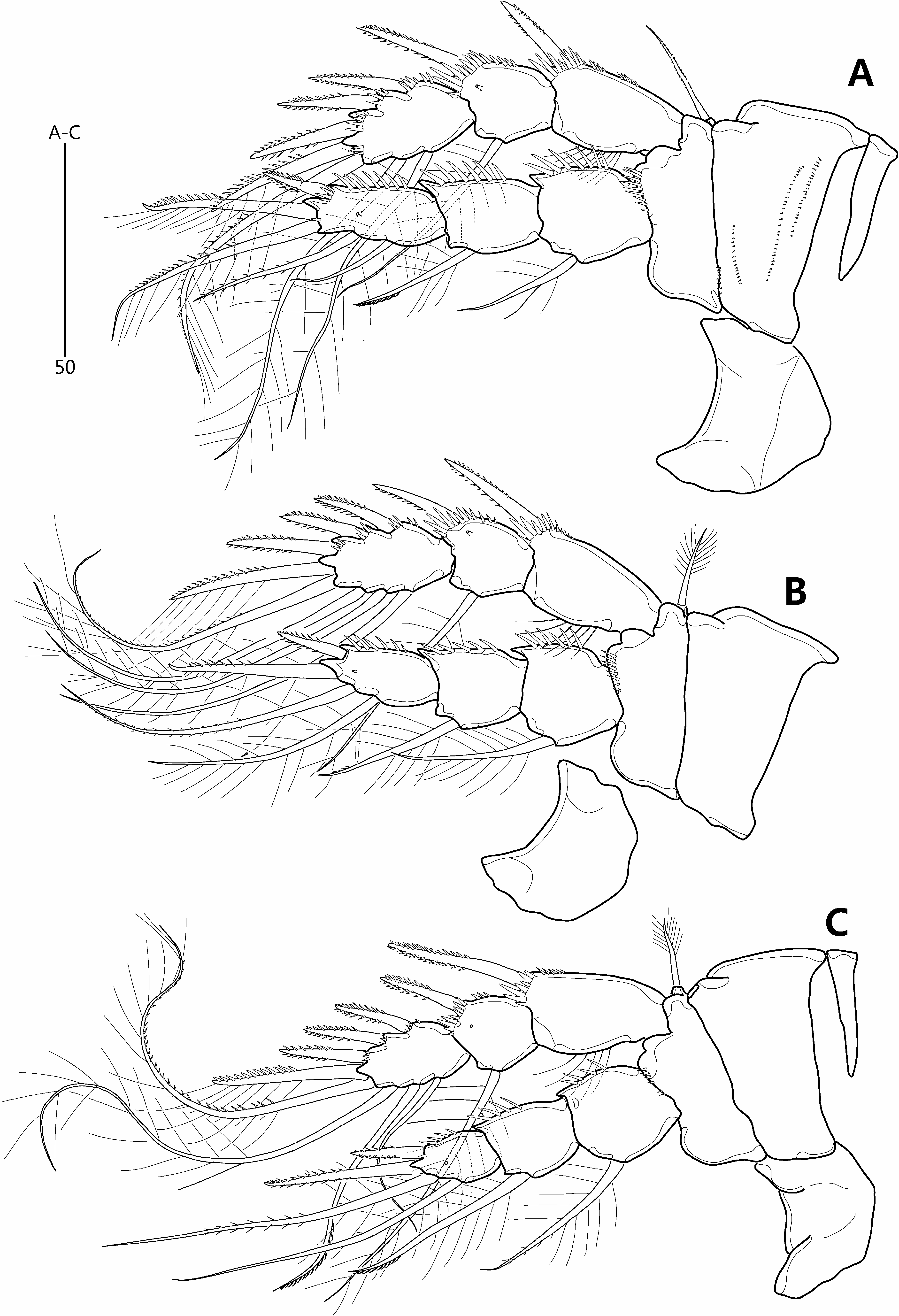
Redescription of female. Total length from anterior margin of rostrum to posterior margin of caudal rami about 615 μm (measured in lateral aspect). Habitus ( Fig. 8A–B View FIGURE 8 ) slightly pyriform, with distinct separation between prosome and urosome; prosome wider than urosome, greatest width measured at P2-bearing somite. Rostrum ( Fig. 8A–C View FIGURE 8 ) large, broad, defined at base, with two apical sensilla and two middorsal pores. Cephalothorax bell-shaped, slightly longer than greatest width, and with sensillar pattern and few pores as figured; P2-bearing somite separated from cephalothorax by large arthrodial membrane. Pedigerous somites gradually tapering posteriorly, ornamented with setules; hyaline frills plain; intersomitic membranes of second to fourth somites discernible in dorsal and lateral aspect. Urosome ( Figs. 8A View FIGURE 8 , 10A View FIGURE 10 ) slightly tapering posteriorly, ornamented with sensilla dorsally and ventrally except for penultimate somite and ventral surface of anal somite. Genital double-somite and abdominal somites 2–3 with multiple transverse rows of fine spinules dorsally. Genital and first abdominal somites completely fused, forming genital double-somite; original segmentation marked externally by slight bilateral constriction and internally by lateral transverse cuticular ribs. Gonopores ( Fig. 10C View FIGURE 10 ) fused, forming common median genital slit; sixth legs (P6) fused, forming single plate with two bare setae either side. Copulatory pore small, located just posterior to genital slit; connected via short copulatory duct with paired anterior seminal receptacles. Posterior margin of penultimate somite with weakly produced dorsal extension, reaching to proximal fourth of anal somite ( Fig. 8A View FIGURE 8 ). Anal somite ( Fig. 8A View FIGURE 8 ) small, cleft medially; with one pair of dorsal sensilla, and minute dorsolateral spinules along posterior margin (not figured); operculum smooth.
Antennule ( Fig. 8D View FIGURE 8 –D’) slender, 7-segmented; with aesthetasc on segments 4 and 7; segment 5 shortest; posterior seta on segment 6 enlarged, densely plumose and bi-articulated at base. Setal armature as follows: 1-[1], 2-[7 + 2 pinnate], 3-[5], 4-[2 + (1 + ae)], 5-[2], 6-[5 + 1 pinnate + 1 plumose], 7-[5 + (1 + ae)]. Each aesthetasc fused basally to adjacent seta.
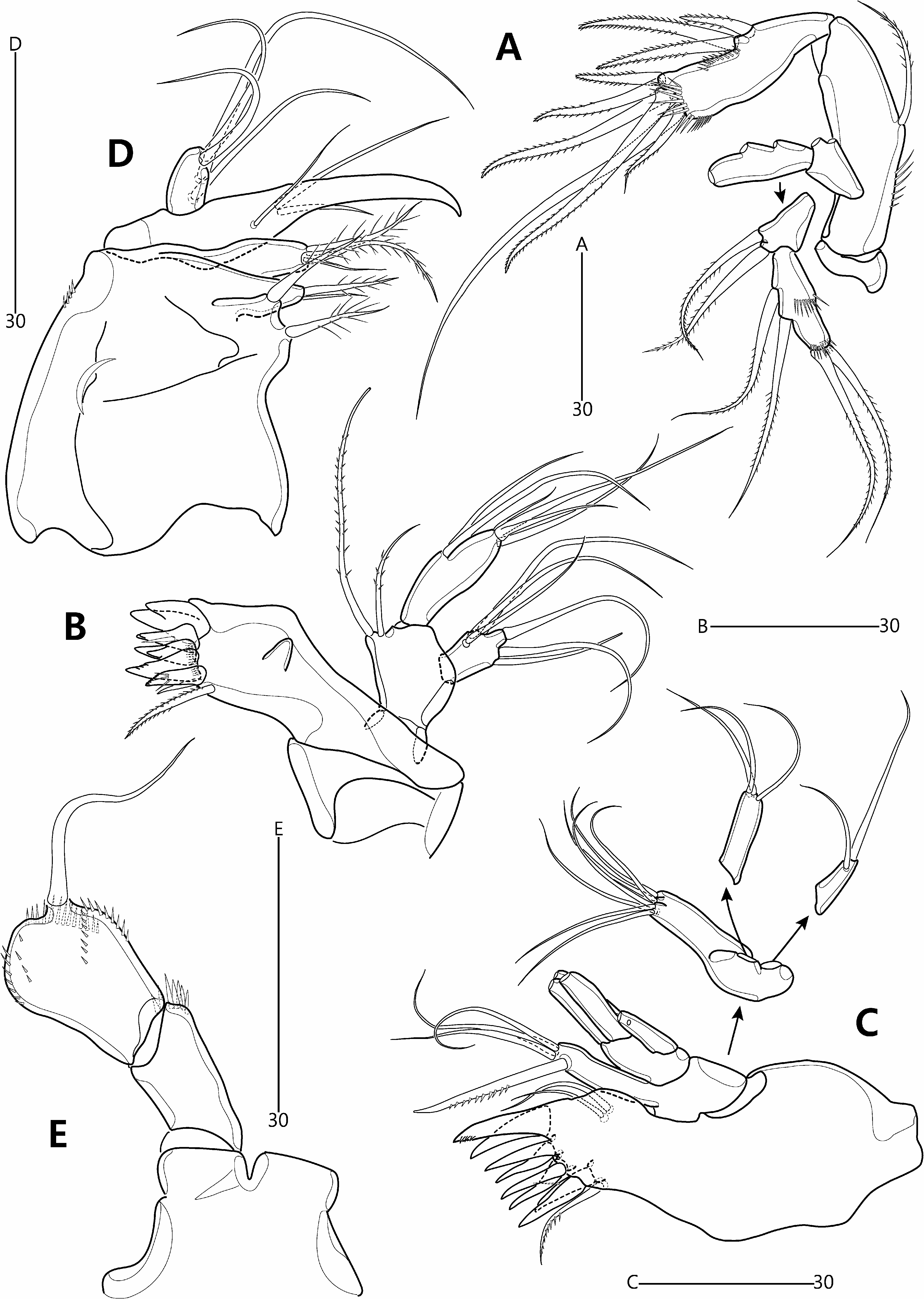
Antenna ( Fig. 9A View FIGURE 9 ) comprising coxa, allobasis and free endopod. Coxa represented by small, unornamented sclerite. Abexopodal margin of allobasis with spinules in basal half and one bipinnate seta halfway down its length. Exopod 2-segmented; exp-1 shortest, with two closely set setae subdistally; exp-2 about 1.5 times as long as exp-1, with two lateral and two apical setae, and two spinule rows; all exopodal setae bipinnate.
Mandible ( Fig. 9B View FIGURE 9 ). Coxa with small pointed process medially; gnathobase comprising five bi- or multicuspidate teeth, one bare spine and one pinnate seta, and ornamented with row of minute spinules. Palp comprising basis and 1-segmented rami. Basis broad, arising from coxal peduncle; with two pinnate setae distally. Exopod small; with three inner, one apical and two outer setae; all exopodal setae naked. Endopod twice as long as exopod; with two lateral, one subapical and three apical setae, two of which fused basally; all endopodal setae bare.
Maxillule ( Fig. 9C View FIGURE 9 ) with well-developed praecoxal arthrite bearing two juxtaposed anterior setae; distal armature consisting of eight spines and one pinnate seta. Coxa with cylindrical endite bearing one unipinnate spine subapically and three naked setae apically. Basal endite with seven naked setae distally. Exopod 1-segmented, with two (sub)apical naked setae. Endopod 1-segmented, longer than exopod, with three apical naked setae.
Maxilla ( Fig. 9D View FIGURE 9 ). Syncoxa armed with few minute spinules along distal outer margin; proximal endite broad, with two plumose setae; middle endite with one plumose and one bare setae apically; distal endite with one bare and two plumose setae. Allobasis drawn out into acutely recurved claw bearing three accessory setae. Endopod 1- segmented, with four setae.
Maxilliped ( Fig. 9E View FIGURE 9 ). Syncoxae arising from common bilobate median pedestal; elongate, with spinules at inner distal corner. Basis expanded in distal half, forming lobate expansion along outer margin; with five rows of minute spinules and one long bare seta near distal inner corner. Endopod not expressed.
P1 ( Fig. 10D View FIGURE 10 ) with wide intercoxal arthrite. Praecoxa small, unornamented. Coxa large, with lobate expansion along outer margin; anterior surface with five rows of minute spinules. Basis smaller than coxa, with bipinnate outer seta and stout, pinnate inner spine; anterior surface with pore and spinules near articulation with endopod and at base of inner spine. Exopod 3-segmented, each segment with spinules along outer margin and outer distal corner; exp-1 longest, with few setules along inner margin; exp-2 about as long as wide; exp-3 slightly longer than exp-2; spines of exp-2 and -3 with spinules along outer margin only. Endopod extending beyond distal margin of exopod, 2-segmented; each segment with spinules along outer and distal margins; enp-1 elongate, not reaching to distal margin of exp-2, unarmed; enp-2 as long as enp-1 but slightly more slender.
P2–P4 ( Fig. 11A–C View FIGURE 11 ). Intercoxal sclerite large, with convex distal margin. Coxa large; anterior surface naked in P3–P4 but with four rows of minute spinules in P2. Basis smaller than coxa, with minutely pinnate (P2) or plumose (P3–P4) outer seta and spinules near articulation with endopod. Exopod 3-segmented, each segment with spinules along outer margin and distal outer corner; exp-1 typically longest, with setules along inner margin; exp-2 about as long wide, with pore on anterior surface; exp-3 longer than exp-2. Endopod 3-segmented; longer than exopod in P2, about as long as in P3, and shorter than in P4; each segment armed with spinules along outer margin; segments of each ramus progressively narrower from exp-1 to -3; enp-3 gradually reducing in size from P2 to P4, with pore on anterior surface. Armature formulae of P1–P4 as follows:
P5 ( Fig. 10E View FIGURE 10 ). Intercoxal sclerite minute. Both rami fused, forming single bilobate plate with deep V-shaped notch between exopodal and endopodal lobes. Endopodal lobe broad and rounded, with four long, evenly spaced, naked setae along distal margin; anterior surface with pore between bases of two outermost setae. Exopodal lobe distinctly smaller and subtriangular; with short outer and long apical seta. Outer basal seta naked with its setophore completely absorbed in outer margin of plate.
Caudal rami ( Figs. 8A–B View FIGURE 8 , 10A–B View FIGURE 10 ) divergent, about twice as long as wide; ventral surface with pore near base of seta II and row of fine spinules along posterior margin; with seven setae: seta I very small, arising halfway down length of outer margin; seta II twice as long as caudal ramus, arising from distal fifth of ramus; setae III and VI long and naked; distal setae IV–V well-developed, pinnate and flexible distally (rat-tail setae sensu Willen (1996)); seta IV shorter than seta V; seta VII plumose, as long as caudal ramus, situated halfway down the ramus near the inner margin.
Male. Unknown.
Remarks. Our Korean specimens agree in virtually every morphological aspect with Pseudomesochra tatianae , originally described from two fjords along the Norwegian west coast ( Drzycimski 1968). The species can readily be identified by the following combination of characters: (1) antennule ♀ with 7-segmented; (2) antennary exopod 2-segmented; (3) extremely reduced maxilliped, lacking expression of the endopod; (4) P1 exp-1 with and enp-1 without an inner seta; (5) P2–P4 exp-3 with three outer spines; and (6) P3–P4 exp-3 with three inner setae. Our Korean female specimens deviate in some minor differences from the original description of P. tatianae such as the presence of caudal ramus seta I, the plumose ornamentation of caudal ramus seta VII, and the spinular ornamentation of the maxillipedal basis and P2 coxa. Such discrepancies are undoubtedly based on erroneous observations of morphological minutiae and are insignificant at the taxonomic level.
Pseudomesochra tatianae belongs to a group of species that share the following character states: antennule ♀ 7-segmented, P1 enp-1 as long as or longer than enp-2, P2–P4 exp-3 with three outer spines, and two, three, three inner setae, respectively, P2–P3 enp-3 with two inner setae; P4 enp-3 with four elements, P 5 ♀ inner exopodal seta as long as or longer than endopodal setae, and caudal ramus about twice as long as wide. This species group includes P. similis Lang, 1936b , P. laptevensis Willen, 1996 and P. tatianae . The latter differs in the morphology of the maxilliped (reduced and endopod not expressed vs endopod represented by small segment bearing seta or short claw) and the length of the inner exopodal seta of P 5 ♀ (about as long as outer endopodal seta vs distinctly longer in P. similis and P. laptevensis ). Pseudomesochra tatianae is also readily distinguished from P. laptevensis by the setal armature of P3–P4 enp-3 (two inner setae in P. tatianae vs only a single seta in P. laptevensis ). The species had not been recorded again since its original description although Easton & Thistle (2016) recently reported six individuals from two deep-sea localities (2,698 –3,247 m) off the western coast of the U.S.A. which they attributed to P. cf. tatianae .
The genus Pseudomesochra assumes a worldwide distribution as demonstrated by the numerous unidentified species that have been reported, including records from the northeastern Pacific off the western coast of the U.S.A. ( Sedlacek 2007; Easton & Thistle 2016), the San Diego Trough ( Eckman & Thistle 1988, 1991), the Clarion Clipperton Fracture zone in the north-east Pacific ( Radziejewska 2014; Radziejewska & Kotliński 2002), the Kuril Trench in the north-west Pacific Ocean ( Kitahashi et al. 2013), the Ryukyu Trench ( Kitahashi et al. 2014), South Andaman ( Jayabarathi 2016), the equatorial North Pacific ( Mullineaux 1987), the Peru Basin ( Willen 1996, 2000), the Magellan Region (Straits of Magellan and Beagle Channel) ( George 1999, 2005), New Zealand ( Webber et al. 2010), the Weddell Sea ( Willen 1996), the deep sea off North Carolina ( Coull 1973), the Gulf of Mexico ( Baguley 2004; Brooks et al. 2009; Degen 2010; Degen et al. 2012; Plum et al. 2015), the Sedlo and Seine Seamounts in the North Atlantic ( Büntzow 2011; George 2013), the Porcupine Seabight ( Gheerardyn 2007), the Svalbard Archipelago ( Kotwicki 2002), the Laptev Sea ( Willen 1996), the Anaximenes Seamount in the eastern Mediterranean Sea ( George et al. 2018) and a marine cave near Marseille ( Janssen et al. 2013). Buhl-Mortensen et al. (2010) found that members of Pseudomesochra represented over 30% of the harpacticoid community found on glass sponge skeletons in the cold-water coral degradation zone. This study was partly based on research by Gheerardyn (2007) (see also Gheerardyn et al. 2009, 2010) who found seven new species in the Porcupine Seabight in the northeast Atlantic, rendering Pseudomesochra one of the most speciose genera in the study area. George et al. (2018) reported ten unnamed species of Pseudomesochra from the Anaximenes Seamount in the eastern Mediterranean Sea. On the lower trench slope of the Kuril Trench ( Kitahashi et al. 2013) and on the abyssal seafloor of the Clarion Clipperton Fracture zone ( Radziejewska & Kotliński 2002) it is the second most abundant genus accounting for over 10% of total harpacticoid density while in the Ryukyu Trench it is the third most abundant genus across the entire bathymetric range ( Kitahashi et al. 2014). Sedlacek (2007) identified six species on the continental slope off central California while Baguley (2004) recorded no less than 23 unidentified species from the deep sea in the Northern Gulf of Mexico. Despite this as yet unexplored species diversity in many deepwater areas of the world’s oceans it is remarkable that a great proportion of the species described so far are only known from the type locality ( Table 3).
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |