
Terranectes parvus, Rahmat & Koretsky & Osborne & Alford, 2017
|
publication ID |
https://doi.org/10.1515/vzoo-2017-0029 |
|
DOI |
https://doi.org/10.5281/zenodo.6461847 |
|
persistent identifier |
https://treatment.plazi.org/id/03A487D1-700D-C37C-4BC5-F9E6FCADF895 |
|
treatment provided by |
Felipe |
|
scientific name |
Terranectes parvus |
| status |
sp. nov. |
Terranectes parvus sp. n. ( figs 4 View Fig , 6 View Fig , 8 View Fig , 9 View Fig , 11 View Fig , 12; tables 1–5 View Table 1 View Table 2 View Table 3 View Table 4 View Table 5 )
urn:lsid:zoobank.org:act:
Etymology. Parvus (Latin, “small”)
Holotype. Femur CMM-V-4709 .
Referred specimens. In addition to the holotype, the following specimens were found in southern Virginia near the North Carolina border: Skull fragment: CMM-V-4679 ; Humeri:
CMM-V-4688, CMM-V-4686 (immature), CMM-V-4687; Ulna: CMM-V-4690 (immature); Innominata: CMM-V-4683, CMM-V-4684; Tibiae: CMM-V-4689 (immature), CMM-V-4778; Fibula: CMM- V-4701; Astragalus: CMM-V-4682; Phalanges (7): CMM-V-4694-4700; Metapodi- als (3): CMM-V-4691-4693.
Type locality. Site #PQ-BH9812, Southampton County, Virginia near the North Carolina border .
Geological age and distribution. Upper Miocene, Chesapeake Group, Eastover Formation (7.0-6.0 Ma) and St. Marys Formation (10.0-8.0 Ma).
Diagnosis. Seals of medium size.
Humerus: deltoid crest extends about ½ of bone length (in contrast to T. magnus ); deltoid tuberosity very pronounced (in contrast to T. magnus ); in lateral view deltoid crest flattened and oval (similar to T. magnus ); lateral epicondyle lower and narrower than the medial, does not reach proximal part of deltoid crest; medial epicondyle extending lower than coronoid fossa; coronoid fossa oval and deep.
Femur: proximal part of greater trochanter wider than the distal; trochanteric fossa deep, wide; head small, seated on distinct lip on short and wide neck; supracondylar fossa shallow, narrow, and elongated (similar to T. magnus ); smallest width of diaphysis shifted proximally (in contrast to T. magnus ). Innominate iliac tuberosity flattened, small and turned anteriorly; alar spine overhangs.
Description—S k u l l: fig. 4 View Fig , F–G. The glenoid fossa measures 16.6 mm anteroposteriorly and 30 mm transversely. Its posterior border forms a very deep, wide postglenoid groove in the tympanic bone. A postglenoid foramen is located at the end of the postglenoid process in this groove, and is not floored by the tympanic bone as in Devinophoca (similar to T. magnus ). The postglenoid process itself is wide ( 24.8 mm) and is located 7.3 mm anterior to the meatal tube.
Laterally ( fig. 4 View Fig , F–G), the bulla is extended as a long tube, with a prominent ventral lip forming the ventral margin of the round external auditory meatus. The rim of the external auditory meatus is separated by a deep but short notch from the mastoid process (as in other carnivores). As in other phocids ( Mitchell and Tedford, 1973), this notch continues as a well-defined groove extending anterolaterally from the stylomastoid foramen along the side of the external auditory meatus. This groove is a synapomorphy of phocids ( Koretsky and Holec, 2002). The pit for the tympanohyal ligament is separated from the stylomastoid foramen (a primitive condition) and is anterolateral to the latter (as in Lontra ; see Muizon, 1982).
In ventral view ( fig. 4 View Fig , F–G), the tympanic bulla is roughly triangular in outline, has a smoothly convex ventral surface, is slightly inflated in its anterior (= ectotympanic) parts, and slopes uniformly to the posterolateral parts. The length of the auditory bulla ( 40.2 mm) is 2.4 times the anteroposterior width of the glenoid fossa, as in other monachines, but in contrast to Devinophoca (3.4 times).The long axis of the bulla is parallel to the midline of the skull. The median lacerate foramen and musculotubular canal with the petrotympanic fissure (= groove) are separated by a thin septum (3.0 mm) above the anteromedial corner of the bulla. The inflated ectotympanic part is bigger than the entotympanic (in contrast to D. claytoni ). Caudally, the entotympanic is more flattened than the ectotympanic along the anteroposterior axis, and is separated from the ectotympanic part of the bulla by a distinct ridge instead of a sulcus. This flatter entotympanic is in contrast to the more inflated entotympanic of Mustelinae and other Phocidae , as noted by Wozencraft (1989).
The medial portion of the entotympanic close to the petrosal bone forms a deep, long fissure around the medial side of the bulla, and the carotid foramen is separated from the posterior lacerate foramen by a thick wall. The carotid canal is partially concealed in the posteromedial wall of the bulla, considerably anterior to the posterior lacerate foramen (primitive conditions according to Tedford, 1977), not reaching the level of the stylomastoid foramen. The posterior opening and the posteromedial process of the carotid canal are visible in ventral view ( Ray, 1976). In T. parvus (similar to L. lenis , but in contrast to D. claytoni ) the carotid canal is parallel to the surface of the basioccipital, but in contrast to D. claytoni its posterior aperture opens in a ventral direction (derived condition, as in other phocines), and has a fully formed margin at its medial side (this is the primitive condition).
The posterior lacerate foramen is impossible to describe because the basioccipital bone is partially broken. However, the septum between the carotid canal and the posterior lacerate foramen is present (in contrast to ursids, otariids, and primitive musteloids; Mitchell and Tedford, 1973; Tedford, 1977; Wolsan, 1993).
The mastoid process is wide, extends far laterally (as in Monachinae ), and forms a pronounced prominence anterolateral to the auditory bulla. The mastoid is so inflated that it is almost the same height as the bulla in lateral view. There is a complete fusion of the posterolateral portion of the meatal lip to the mastoid process as in other carnivores, but the deep groove is present anteromedially. The continuous crest extending from the mastoid process over the external auditory meatus to the postglenoid process is very well developed.
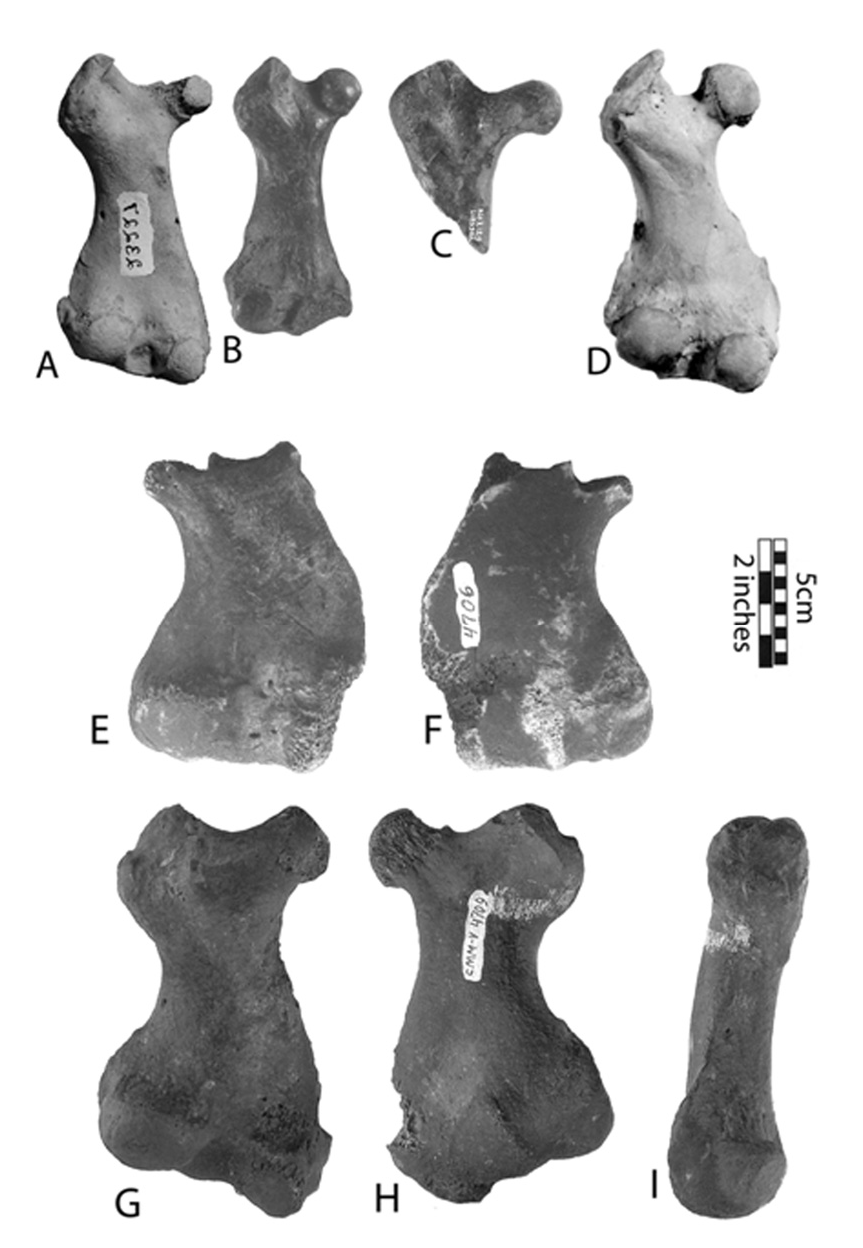
Humerus: fig. 6 View Fig , H–J; table 1 View Table 1 . The deltoid crest extends about half the length of the bone, and smoothly descends to the condyles as a flat, almost invisible ridge. The deltoid tuberosity is large and located at the distal end of the deltoid crest. The lateral epicondyle is longer and narrower than the medial, and does not reach the distal part of the deltoid crest. The medial epicondyle is flattened, extending lower than the coronoid fossa. The entepicondylar foramen is absent. The coronoid fossa is deep, oval, extends further proximally than the lateral epicondyle, and ends at the same level as the medial epicondyle. The olecranon fossa is very shallow.
Innominate: figs 7 View Fig , 8 View Fig , B, H, K, L; table 3 View Table 3 . As in other Monachinae and Cystophorinae, the ilium is thick and the iliac crest is not everted and not excavated on its ventral surface. The iliopectineal eminenceis well pronounced (as in T. magnus ), and situated higher than the proximal border of the acetabular fossa. The iliac tuberosity is flattened, small and turned anteriorly (in contrast to T. magnus ). The greater ischial notch is slightly concave, almost straight. A shallow depression (fossa) for the gluteus medius m. is located on the lateral aspect of the ilial wing. The alar spine protrudes very far. The edges of the acetabular fossa are raised above the plane surface of the bone. The acetabulum is circular with a deeply marked cotyloid notch. The ischium is flattened, wide and thin. The ischial spine is small, elongated and well developed for attachment of the biceps femoris m. The pubis is not preserved.
Femur: fig. 9 View Fig , G–I; table 4 View Table 4 . The greater trochanter extends proximally slightly higher than the head; its proximal part is wider than the distal. The trochanteric fossa is deep, wide, and opens distally, reaching the distal border of the head. The flat and wide intertrochanteric line disappears at the middle of the diaphysis, which serves for insertion of the iliopsoas m. The femoral head is small, relative to the bone’s mass, and is seated on a very short and thick neck. Between the head and neck is a distinct lip. The smallest width of the diaphysis is shifted toward the proximal end of the bone. The supracondylar fossa is located above the lateral condyle and is shallow, narrow, and elongated. The condyles are unequal in size.
Tibia: fig. 10 View Fig , G–I; table 5 View Table 5 . The two condyles are strongly concave in their centers, shortened, small, and oval. The intercondyloid eminences are large and rise above the two lateral, weak borders of the condyles. The popliteal notch is deep, wide, and well-marked. The tibial crest is rounded in a dorsomedial direction. On the ventral side of the tibia, the tibial tuberosity is flattened and well-marked.
Ulna: fig. 11 View Fig , D–F; table 2 View Table 2 . The medial surface of the bone is concave. The olecranon is short and thin, gradually connecting to the proximal half of the bone. On the lateral surface, is a very visible prominence of the rugosity for insertion of the brachialis m. A protuberance is present for origin of the abductor pollicis longus m. A sharp crest is located caudal to the articular surface. On the bone’s radial aspect, the coronoid process protrudes only slightly forward over the radial notch, which is deep and wide. The interosseous crest is sharp, forming a prominence. The head is not preserved.






No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.