
Ophiacantha scissionis, Lee & Stöhr & Bae & Shin, 2019
|
publication ID |
https://doi.org/ 10.6620/ZS.2019.58-08 |
|
DOI |
https://doi.org/10.5281/zenodo.8055812 |
|
persistent identifier |
https://treatment.plazi.org/id/03A2CF0B-FFE8-8570-FC6F-FEEFEDFD0F74 |
|
treatment provided by |
Valdenar |
|
scientific name |
Ophiacantha scissionis |
| status |
sp. nov. |
Ophiacantha scissionis View in CoL View at ENA sp. nov.
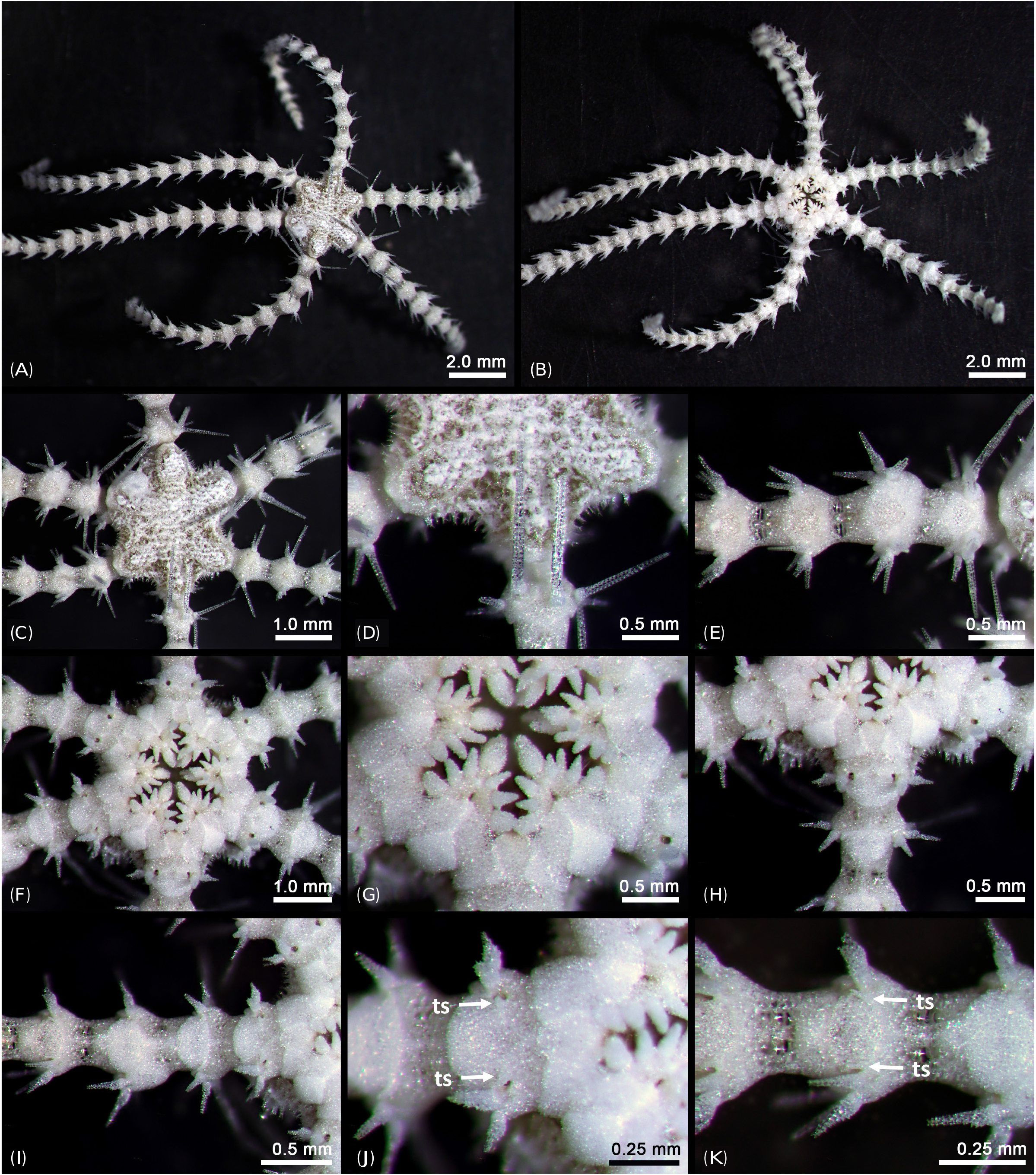
( Figs. 2–11 View Fig View Fig View Fig View Fig View Fig View Fig View Fig View Fig View Fig View Fig ) urn:lsid:zoobank.org:act:99974EF7-D80E-4122-8E6F-268B977F50C5
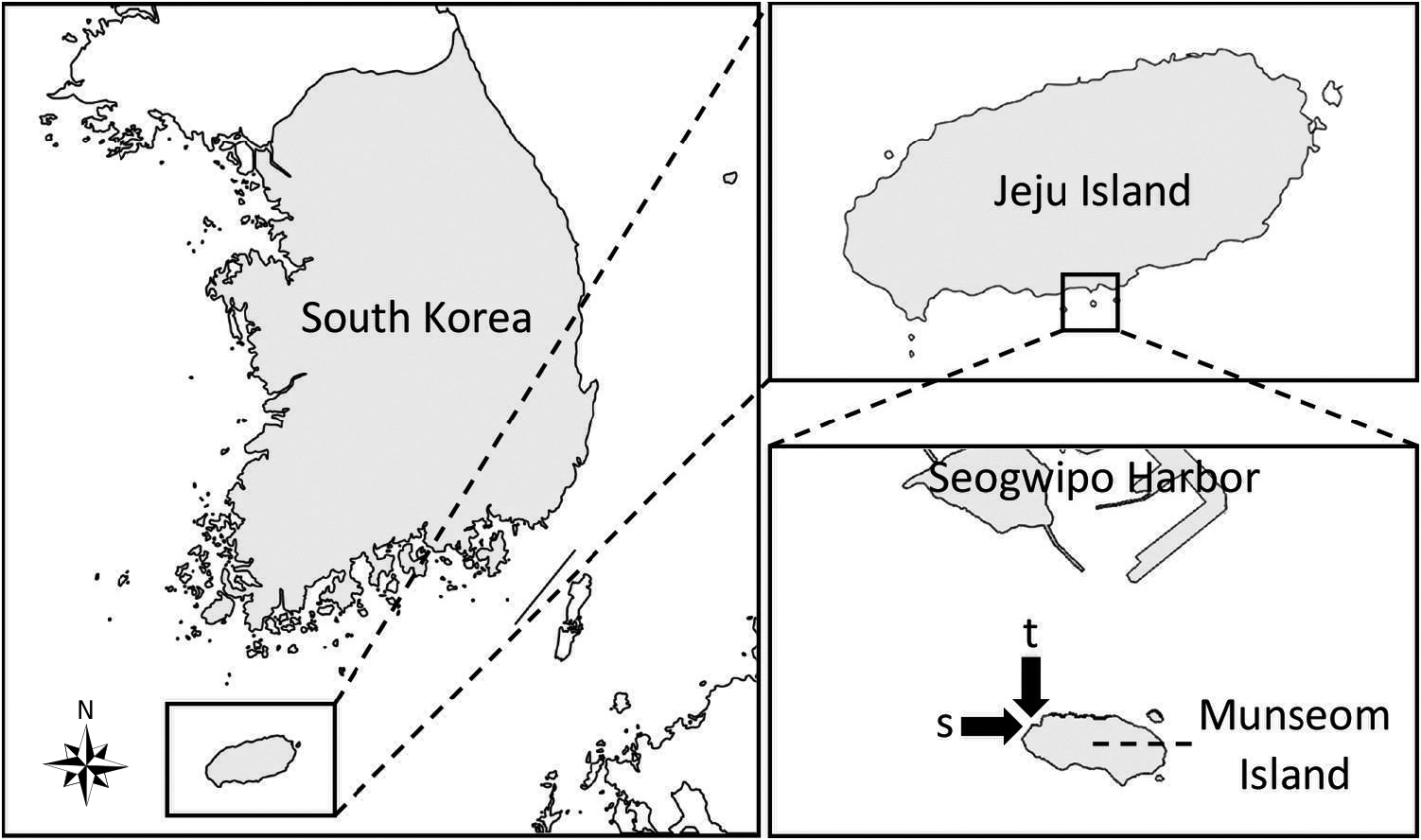
Material examined: Holotype: MERBK-O-21898, Munseom Island , southern Jeju-do, South Korea (33°13'38.2"N, 126°33'47.4"E), depth 35.7 m, water temperature 19.3°C, 27 November 2018, SCUBA diving, T. Lee ( Fig. 2 View Fig ) GoogleMaps . Paratype 1: MERBK-O-21899, Munseom Island, southern Jeju-do, South Korea (33°13'38.2"N, 126°33'47.4"E), depth 31.5 m, water temperature 19.3°C, 27 November 2018, SCUBA diving, T. Lee. Paratype 2: MERBK-O-21900, Munseom Island, southern Jeju-do, South Korea (33°13'38.7"N, 126°33'47.3"E), depth 32.8 m, water temperature 19.3°C, 28 November 2018, SCUBA diving, T. Lee. 29 specimens (MERBK-O-21901), Munseom Island, southern Jeju-do, South Korea (33°13'38.2"N, 126°33'47.4"E), depth 35.7 m, water temperature 19.3°C, 27 November 2018, SCUBA diving, T. Lee. 43 specimens (MERBK-O-21902), Munseom Island, southern Jeju-do, South Korea (33°13'38.7"N, 126°33'47.3"E), depth 32.8 m, water temperature 19.3°C, 28 November 2018, SCUBA diving, T. Lee.
Type locality: Munseom Island, Jeju-do, South Korea, 33°13'38.2"N, 126°33'45.4"E, 35.7 m ( Fig. 1 View Fig ).
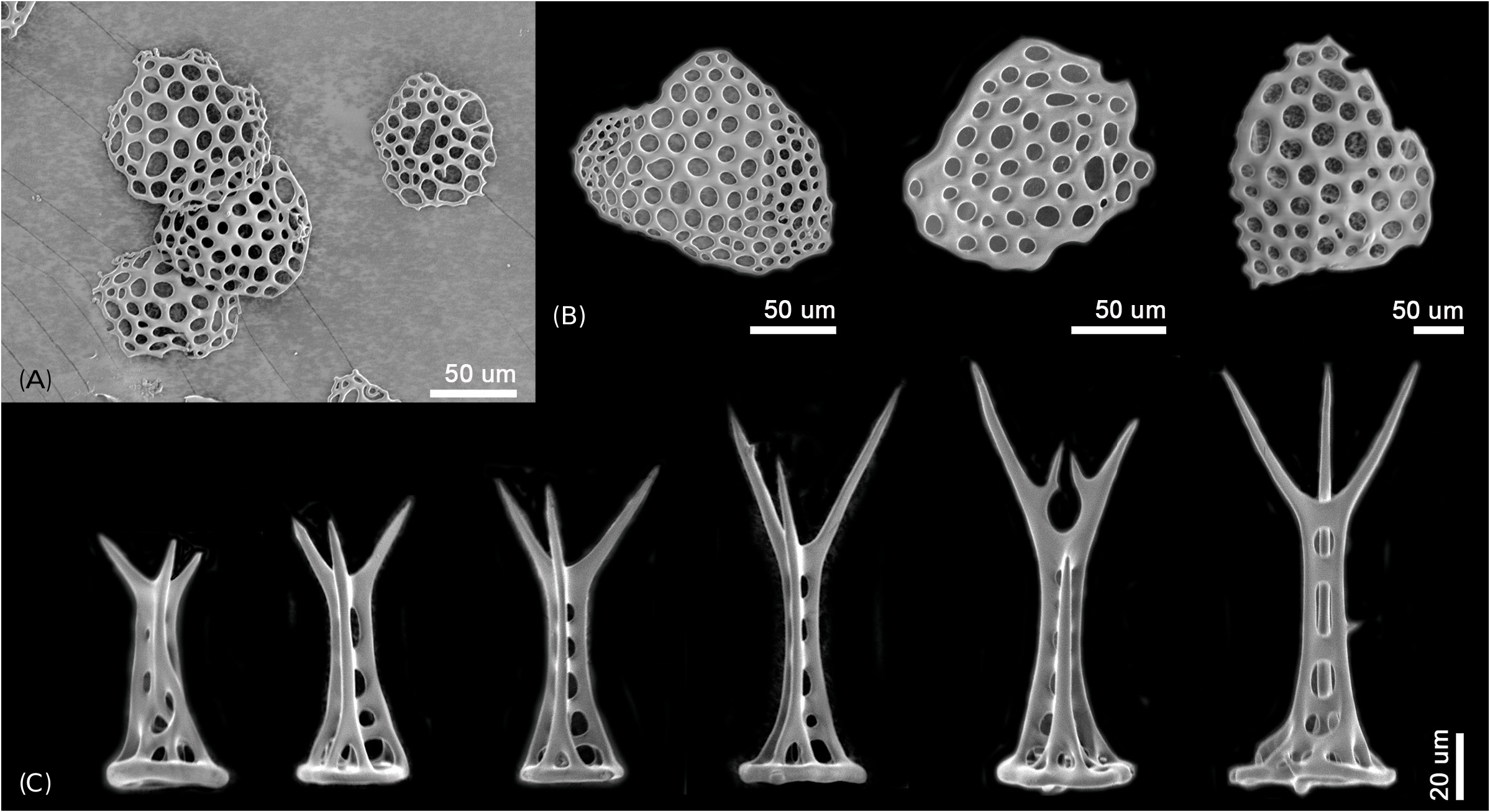
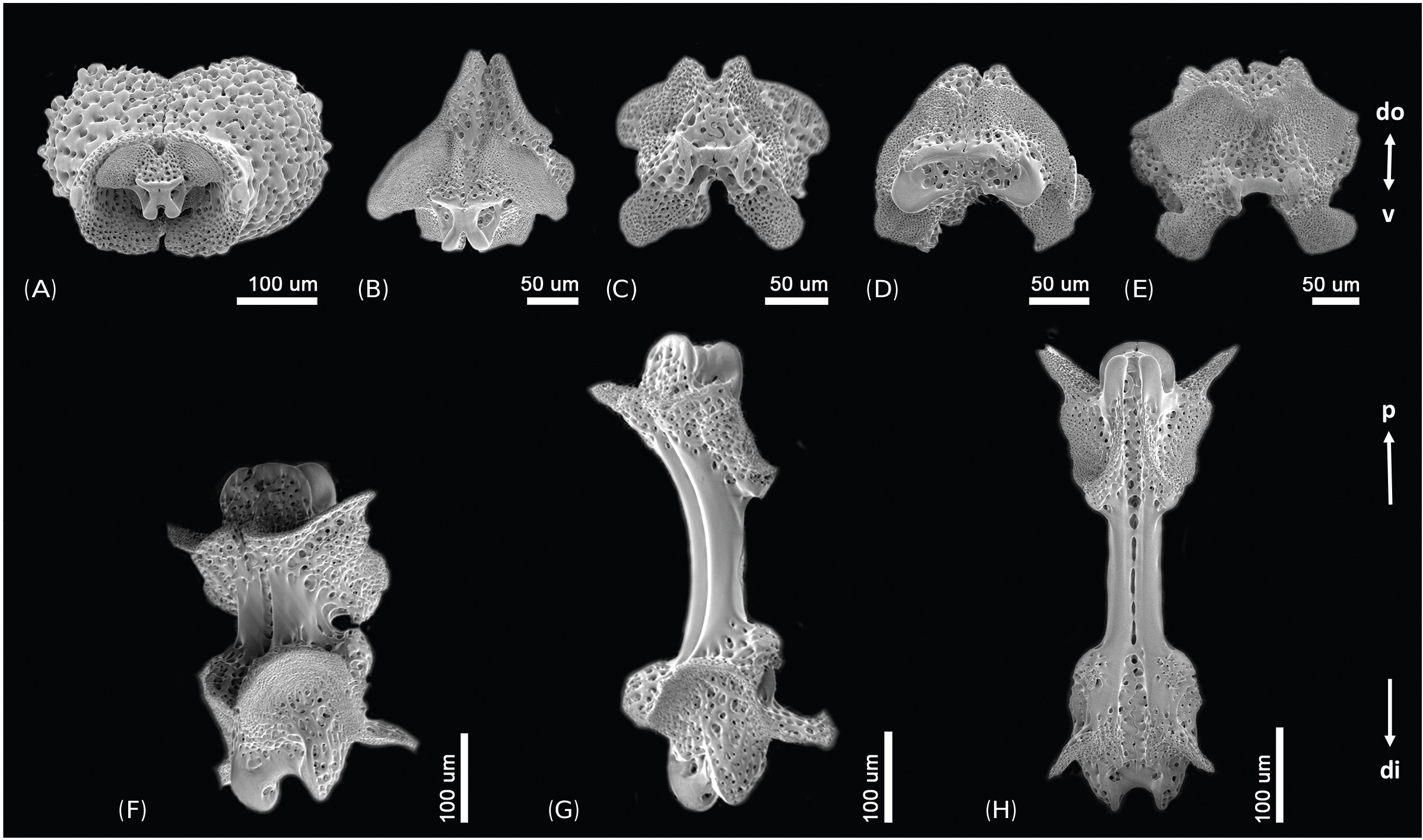
Description of the holotype: Disk diameter 2.0 mm, moderately high, constricted interradially, six arms of almost same size, length of longest arm 9.8 mm ( Figs. 2A, B View Fig ). Arms distinctly moniliform ( Figs. 2A, B, E View Fig ). Disk covered with small, perforated, overlapping scales concealed by thin skin. Small spines densely cover aboral side of disk ( Figs. 2C, D View Fig ), distal part split into three sharp thorns, pillar of spine perforated, basal part usually circular.
Oral shields longer than wide ( Figs. 2F, G View Fig ), with straight or slightly concave margins. Adoral shields longer than wide ( Fig. 2G View Fig ), almost 2.5 times as long as wide. Jaw as wide as long ( Fig. 2G View Fig ), with one blunt and serrated ventralmost tooth. Oral papillae interpreted as proximalmost conical, pointed infradental papilla, 1–2 similar pointed lateral oral papilla(e), and distalmost a larger adoral shield spine with blunt tip ( Figs. 2G, H View Fig ).
First dorsal arm plate as wide as long with obtuse proximal angle and convex distal border ( Fig. 2E View Fig ). Succeeding plates separated, similar to first dorsal arm plate and guitar pick-shaped, wider than long. Lateral arm plate twice as long as wide, with distinctly concave proximal lateral side and distal side more convex than proximal side ( Figs. 2E, I, K View Fig ). Arm spines four proximally, straight, tapered to tip, slightly serrated. Uppermost arm spine longest, reaching to next dorsal arm plate, three times longer than other arm spines at same lateral arm plate ( Fig. 2D View Fig ). After third lateral arm plate, uppermost arm spine rapidly shortening, similar to other arm spines at same lateral arm plate. One tentacle scale across entire arm, proximally a short and obtuse scale ( Fig. 2J View Fig ), further out a small spine-like scale ( Fig. 2K View Fig ). Color in life brownish-gray.
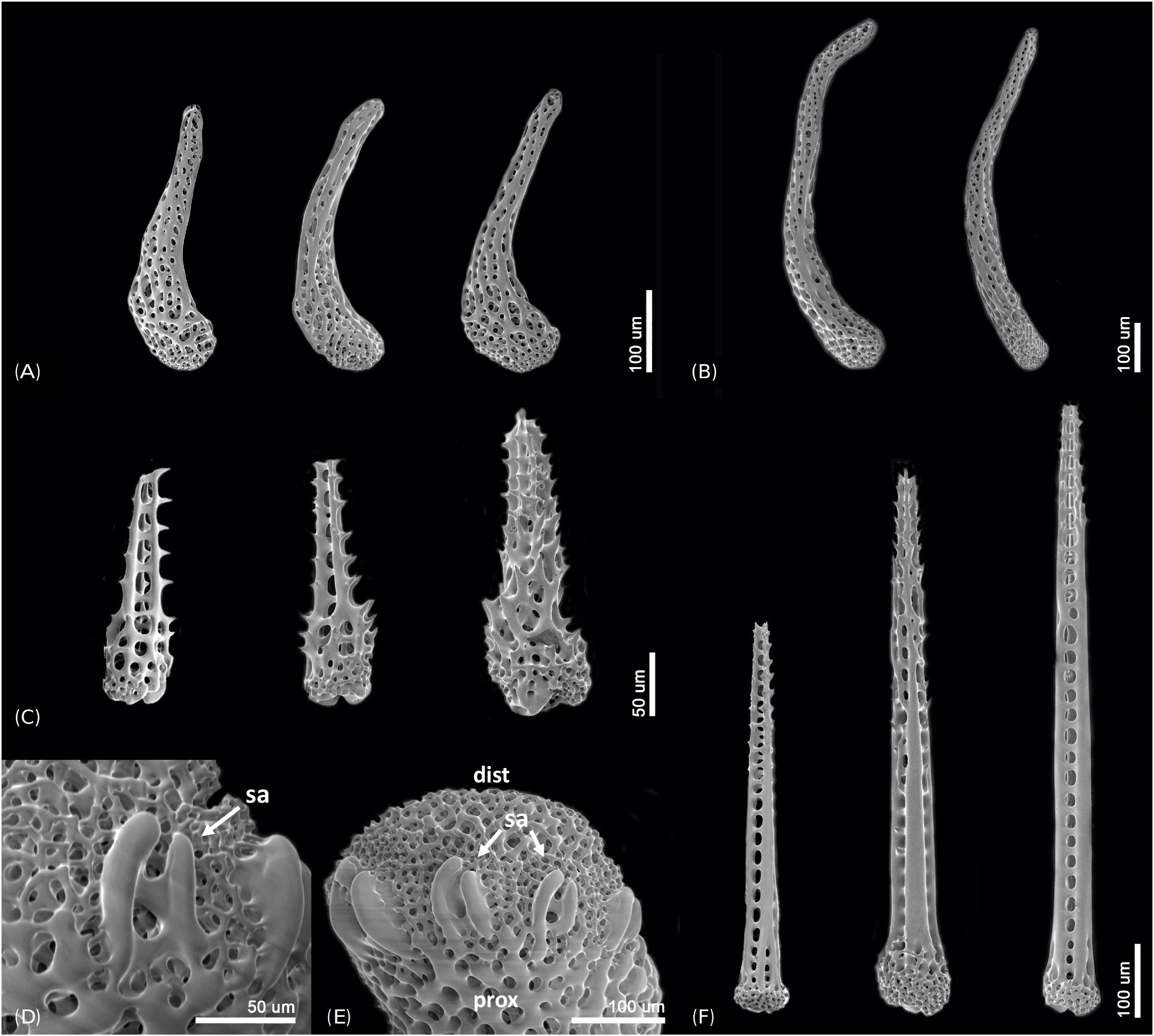
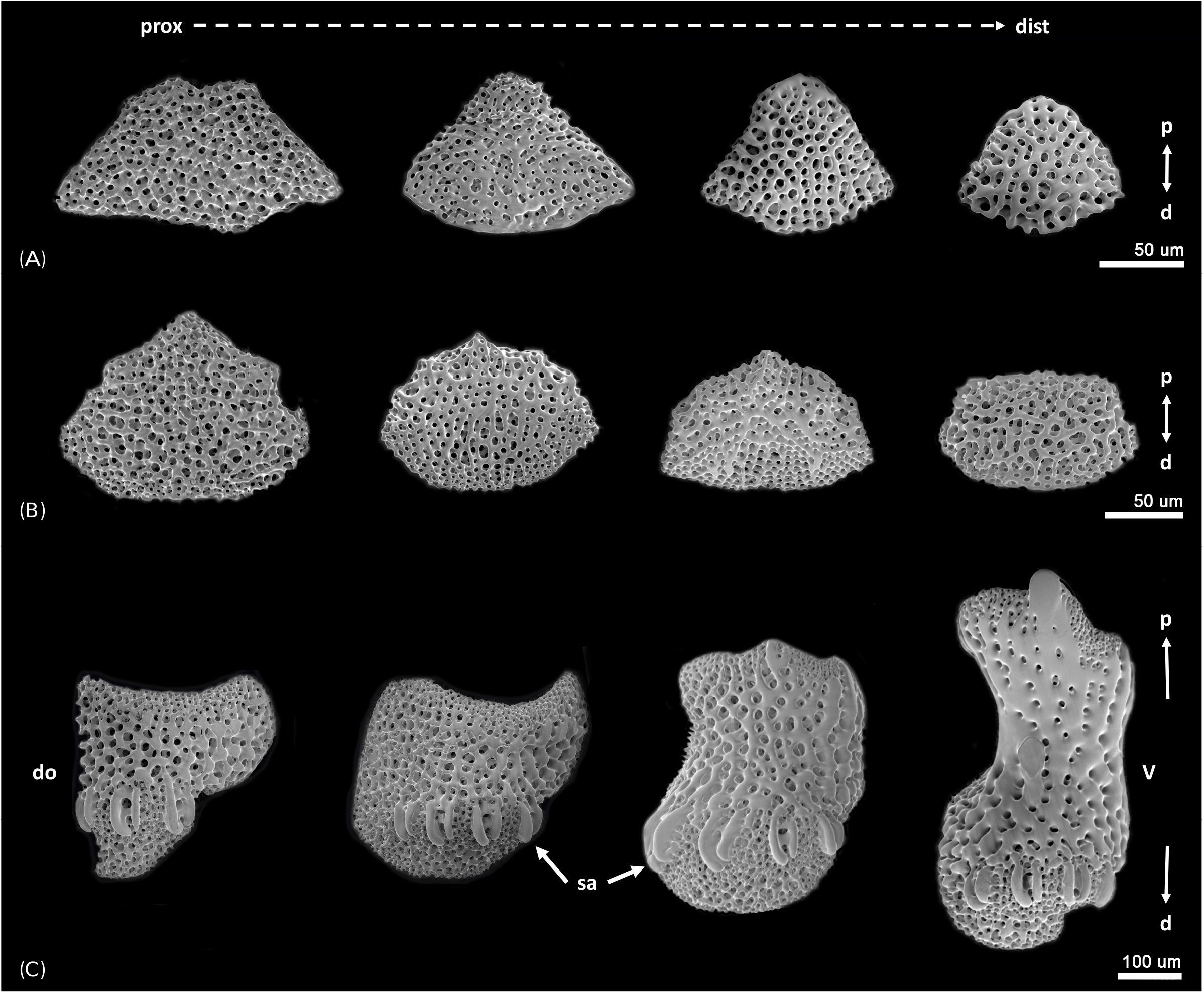
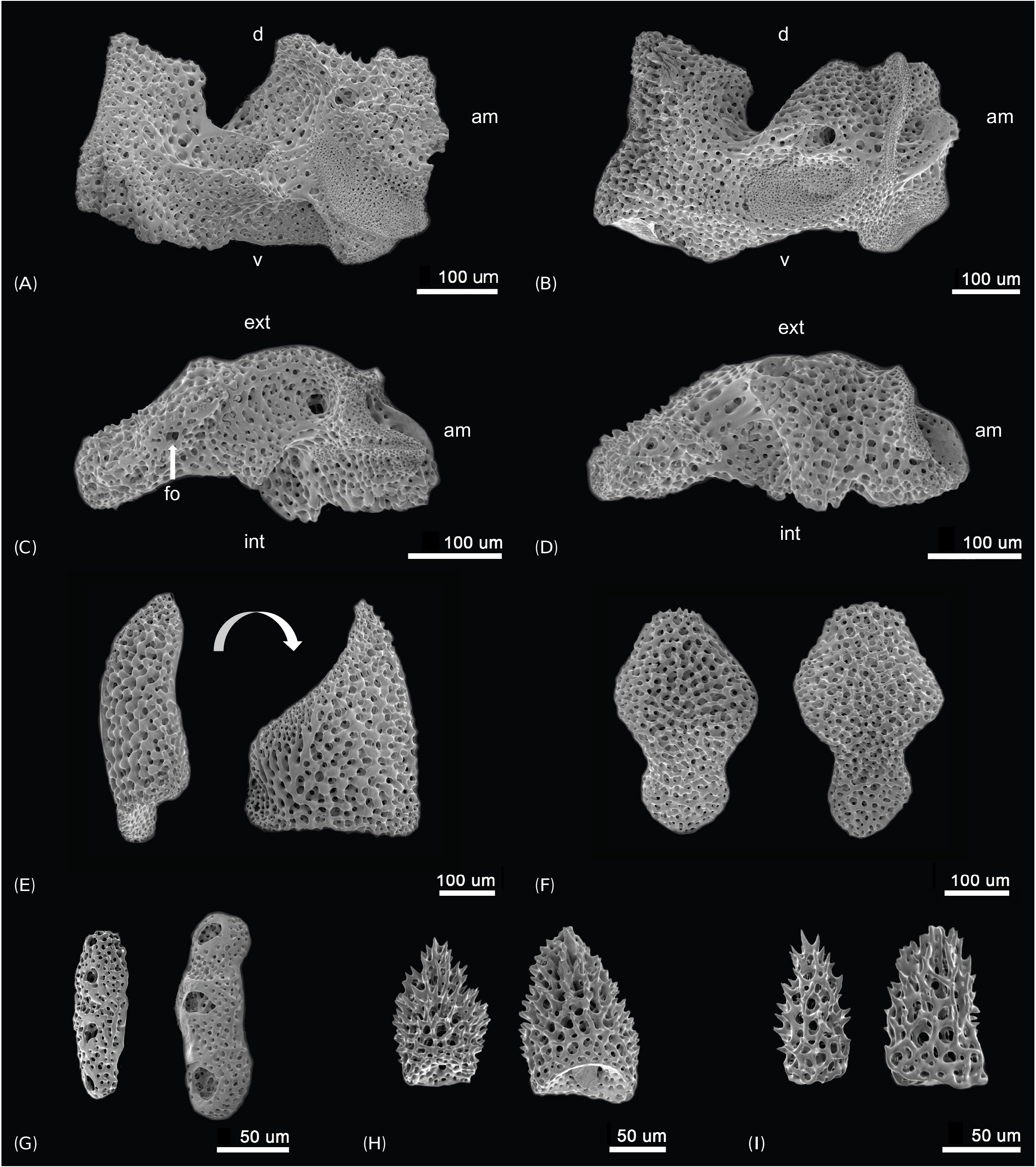
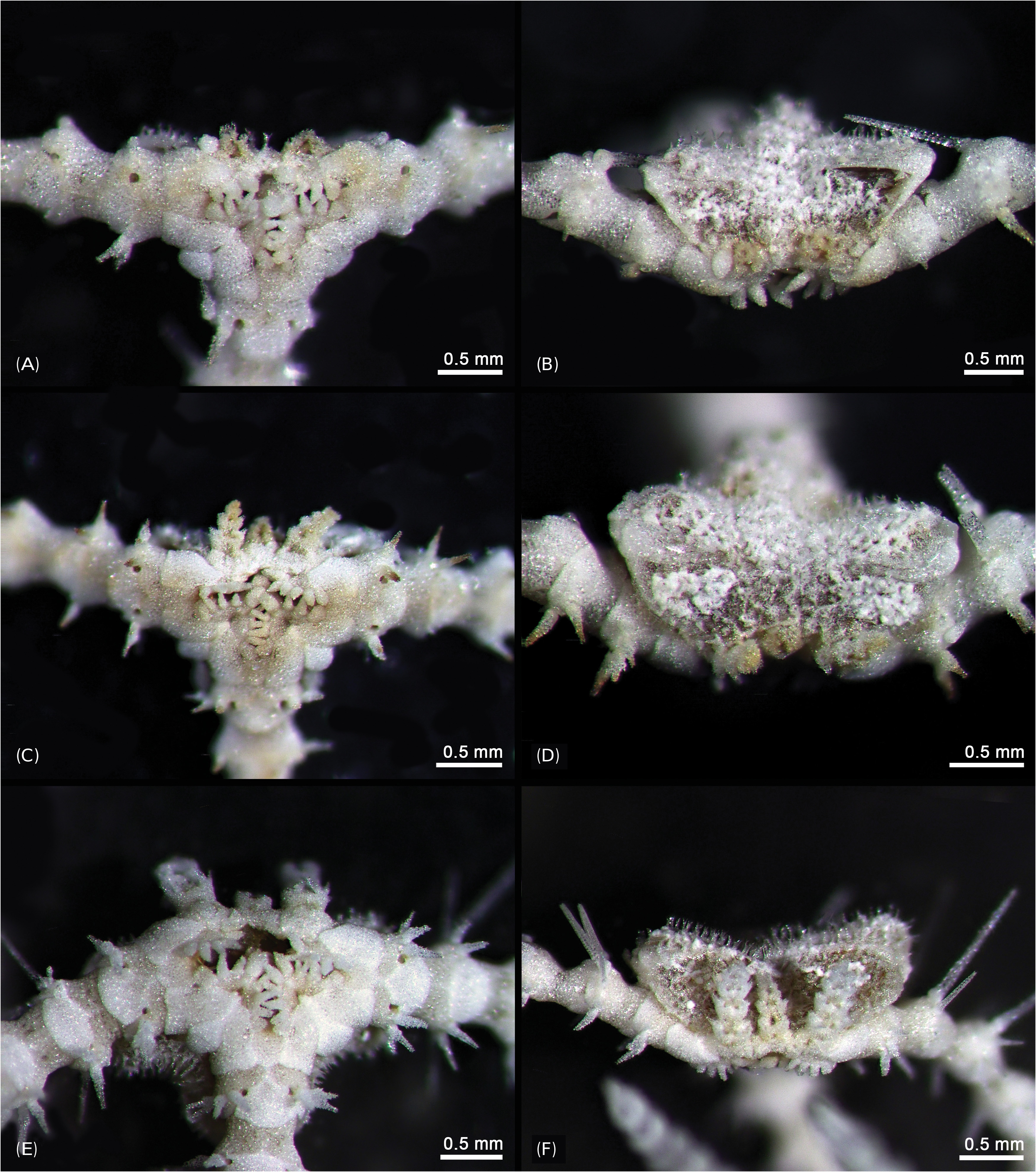
Description of the paratypes: Paratype 1 and 2 are six-armed specimens. The paratypes and other 72 specimens share the same morphological characteristics with the holotype, except for number of arms. Disk diameter 1.3–2.2 mm in two paratypes and 72 specimens, usually with six arms, rarely five and only one specimen with seven arms. Arm spines four to five proximally. Disk scales perforated irregular circles, covered with thin skin ( Figs. 3A, B View Fig ). Disk spines usually trifid but sometimes branched tips present ( Fig. 3C View Fig ). Genital plates small, narrow, tapered to tip and boomerang-shaped ( Fig. 4A View Fig ). Radial shield narrow, long and bent, almost a third of disk diameter long ( Fig. 4B View Fig ). Arm spines usually four proximally, dorsalmost longest ( Fig. 4F View Fig ), rapidly getting shorter ventralwards ( Fig. 4C View Fig ). Dorsal arm spines with serrated tips ( Fig. 4F View Fig ), ventral arm spines more coarsely serrated ( Fig. 4C View Fig ). Arm spine articulation not of typical ophiacanthid form, strikingly similar to amphilepidid form with two parallel lobes (dorsal lobe strongly bent), only weakly connected at their proximal ends, lacking a sigmoidal fold ( Figs. 4D, E View Fig ). Dorsal and ventral arm plates getting shorter and rounder from proximal to distal ( Figs. 5A, B View Fig ). Lateral plate and vertebrae getting longer from proximal to distal ( Figs. 5C, F–H View Fig ). Vertebrae elongated, with zygospondylous articulation ( Fig. 6 View Fig ). Adoral muscle flanges larger than oral muscle flanges at proximal side ( Figs. 6A, B View Fig ) and oral muscle flanges of distal side wider than proximal side ( Fig. 6C View Fig ). Muscle flanges of proximal side slightly wider than distal side ( Fig. 6F View Fig ). Oral plates (half-jaws) elongated, almost twice as long as high ( Figs. 7 View Fig A-D), and adradial muscle attachment area small ( Fig. 7B View Fig ). On ventral surface of vertebra, a flat area of smooth and less perforated stereom with a conspicuous pore, interpreted as fossa and articulation surface for oral papillae ( Fig. 7C View Fig ). Adoral shields longer than wide and larger than oral shield ( Fig. 7E View Fig ). Oral shields longer than wide and constricted at middle part ( Fig. 7F View Fig ). Dental plate small, convex, elongate, with three to four round tooth sockets with low smooth borders, not perforating the plate ( Fig. 7G View Fig ). Ventralmost tooth largest, second tooth smaller, both lanceolate in shape and larger than oral papillae ( Fig. 7H View Fig ). Infradental papilla and lateral oral papilla small, similar, slightly conical, and adoral shield spine larger than others, with blunt tip ( Fig. 7I View Fig ). First tentacle pore larger than others ( Fig. 8 View Fig ). Tentacle scales getting longer and sharper from proximal to distal ( Fig. 8 View Fig ).
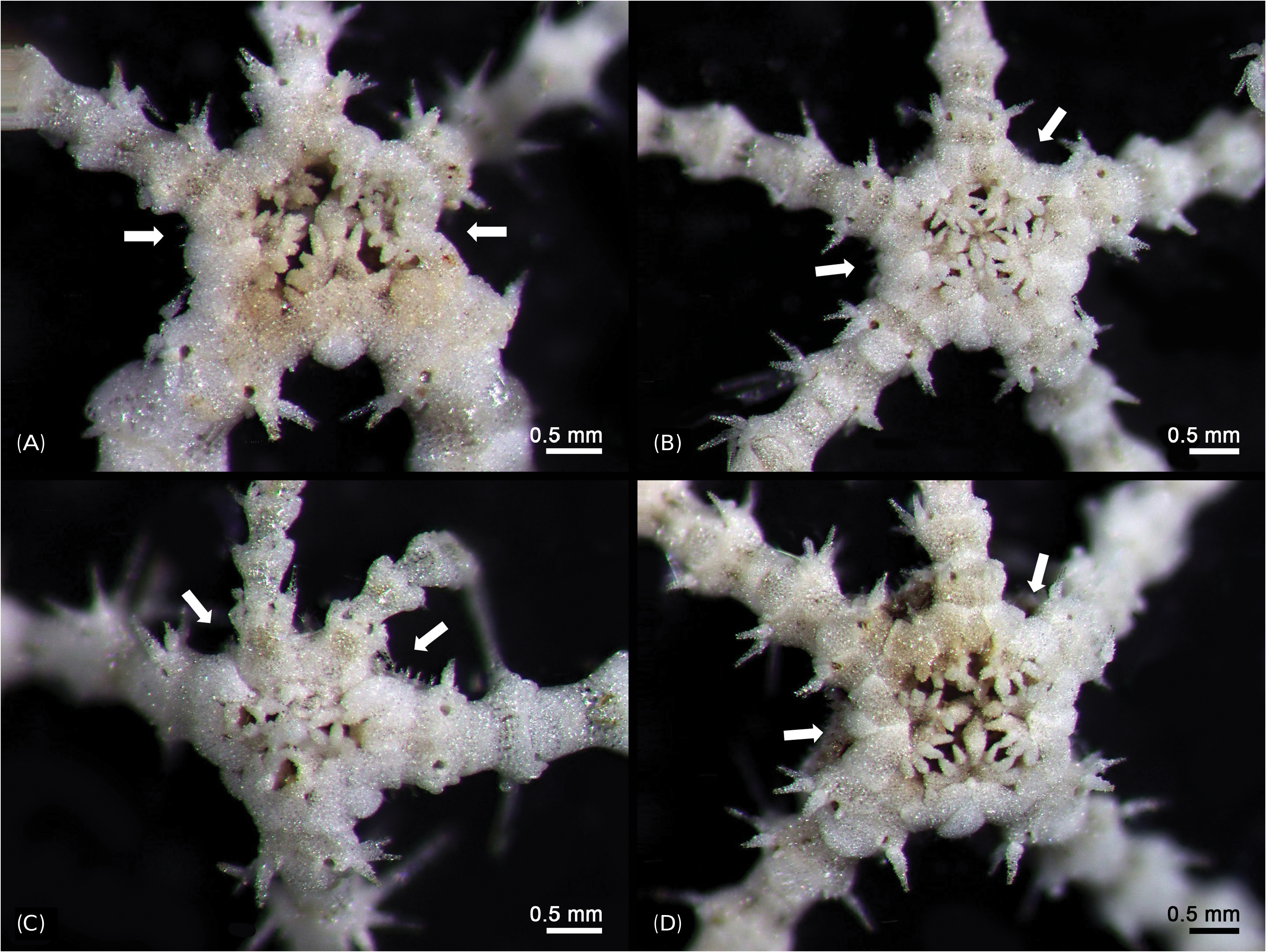
Habitat and ecology: All specimens of O. scissionis sp. nov. were found on a bryozoan, Bantariella bocki ( Silén, 1942) . This bryozoan is distributed from southern Japan (Bonin Islands) to Jeju Island, South Korea; it has been suggested that the species is spreading due to increasing water temperatures ( Seo et al. 2009). The Bonin Islands are located at similar latitude as Taiwan and at the boundary between subtropical and tropical waters ( Seo et al. 2009). The sampling location, Munseom Island in Southern Jeju Island, is affected by a warm current (the Kuroshio current) all year round and the water temperature in the waters surrounding Munseom Island ranges from 16 to 26°C over a year ( Ko et al. 2008). In deep water off Munseom Island, we observed that bryozoans covered black coral ( Myriopathes japonica ( Brook, 1889) and M. lata (Siberfeld, 1909)) . Specimens of O. scissionis sp. nov. were not found on corals, they inhabited only the bryozoan B. bocki . It is at present unknown if the ophiuroids obligately occur on these bryozoans and are spreading together with them. Several species of ophiuroid are known to live epizoic on black coral, e.g., species of Ophiocanops ( Stöhr et al. 2008) and Astrogymnotes ( Baker et al. 2001) . Little is known about these associations, but the corals may provide raised vantage points from which the ophiuroids can collect food particles in the water column, and the brittle stars may hide from predators among the branches of the corals. Although O. scissionis sp. nov. seems not to be associated with the corals themselves, it probably indirectly benefits from them. Perhaps the filter feeding current created by bryozoans also transports food to the ophiuroids. More research is needed to understand these relationships.
Remarks: Most specimens of O. scissionis sp. nov. have an incomplete disk and six arms of different size. Three of the six arms are complete and long but the others seem to be regenerating, as does the disk. This species appears to reproduce asexually by fission. The series of oral papillae was interpreted based on their position on the jaw edge compared to Ophiacantha bidentata (Bruzelius, 1805) , as shown by Hendler (2018), because no developmental series was available to trace the origin of each papilla. We agree that the term apical papilla has been applied to a variety of nonhomologous structures and should be abandoned. The term infradental papilla has been redefined by Hendler (2018) to the proximalmost papilla on the oral plate, not on the dental plate as was long believed. The fossae and articulation surfaces of the oral papillae on the oral plate appear not well defined in O. scissionis sp. nov., except for the middle one. This may perhaps be a sign of incomplete regeneration in the selected specimen.
Etymology: The specific name is derived from the Latin term scissio, meaning split.


| T |
Tavera, Department of Geology and Geophysics |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
SuperOrder |
Ophintegrida |
|
Order |
|
|
SubOrder |
Ophiacanthina |
|
Family |
|
|
Genus |