
ACARI
|
publication ID |
https://doi.org/ 10.1206/3908.1 |
|
DOI |
https://doi.org/10.5281/zenodo.5057093 |
|
persistent identifier |
https://treatment.plazi.org/id/03A187BF-FF84-2F30-A846-470F53AEFB31 |
|
treatment provided by |
Felipe |
|
scientific name |
ACARI |
| status |
|
ACARI (MITES)
Figures 5H View FIG , 11A View FIG
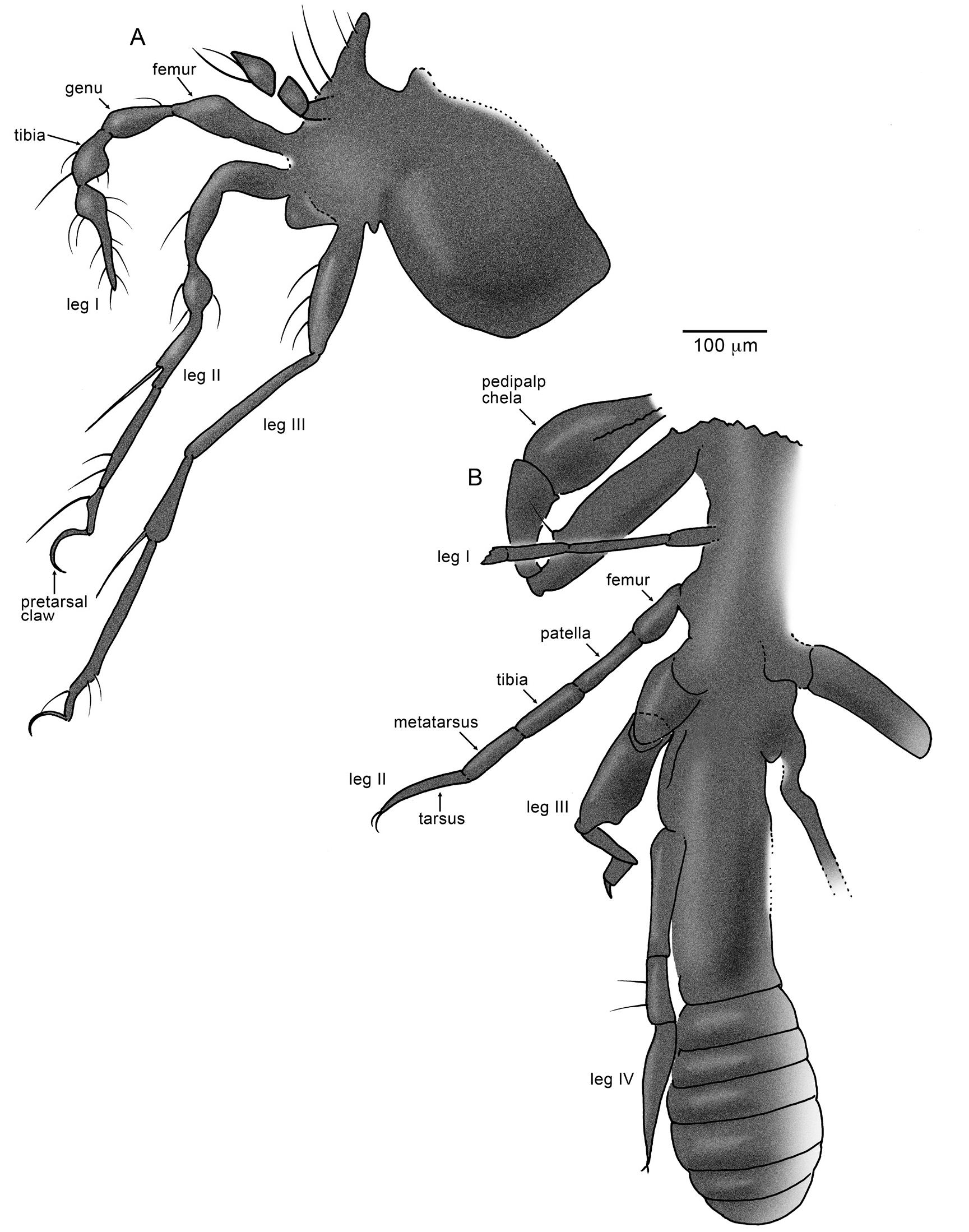
AMNH WH-6: A small piece containing most of the remains of an oribatid mite, similar in overall structure to the family Damaeidae (fig. 11A), which consists of fewer than 100 living species in 14 genera, primarily as mycophagous and algophagous inhabitants of leaf litter and subcortical microhabitats of temperate and boreal forests. Damaeidae occur in Eocene amber from the Baltic region and Rovno, Ukraine ( Weitschat and Wichard, 2010; Perkovsky et al., 2010). The mite is near the corner of the amber piece, with the appendages of one side lost or completely obscured; the piece also contains dark layers from various resin flows as well as particulate plant matter. Body length (without appendages) is approximately 400 μm, with a slight constriction between the prodorsum and notogaster; legs are long and slender, length of the longest is 550 μm. The mite is dark and opaque, generally obscuring many of the setae, sensilla, and cuticular details except those visible at margins. Anterior-most appendage (pedipalps) (only one of a pair observable), with three short, stout podites, apical one pointed, with two long, fine solenidia. A pair of long, stiff solenidia occurs at the anterior end of the prodorsum. Structure of the legs is very distinctive and quite similar to that of the living family Damaeidae , in particular the “moniliform” legs (although this habitus also occurs in oribatid superfamilies closely related to Damaeoidea). Leg I has bulbous portions of the femur, genu, tibia, and at the base of the tela + basitarsus. The only bulbous portion of leg II is on what is either the genu or tibia (boundaries between podites are barely visible). The apices of tibiae in legs II and III each have a long, stiff solenidium that is nearly equal in length to that of its respective podite. Pretarsal claws are long, slender, and sickle shaped.
AMNH LC-D1: A piece of amber containing a complete mite (~280 μm body length), which is moderately well preserved (fig. 5H). Cuticle of the notogaster and prodorsum is cracked and slightly disintegrated, precluding observation of most of the chaetotaxy, glands, and sensilla, though a pair of thick, plumose trichobothria/bothridial setae is visible, one at each posterolateral corner of the prodorsum. While identification of the mite in the Oribatida is certain, a more detailed identification will be challenging. The mite is rather generalized in structure, lacking specialized (e.g., plumose) setae and obvious cuticular microstructure (e.g., reticulations). The gnathosoma is well integrated and conical, laterally with a pair of projecting solenidia; legs are relatively short, pair I with an elongate solenidium dorsoapically on what appears to be the genu; all pretarsal claws are long, slender, and hooked.
The Oribatida is a highly diverse, speciose group of mites comprised of some 9000 living species in 172 families, largely inhabitants of soils, leaf litter, and moss (Norton and BehanPelletier, 2009). The fossil record of the group is ancient and diverse, beginning with unambiguous cuticular remains from the Devonian that are preserved in microscopic detail ( Norton et al., 1988). Oribatids have even been implicated in the processing of plant detritus from Carboniferous swamps ( Labandeira et al., 1997). Their fossil record in amber from the Cretaceous and Cenozoic is excellent ( Dunlop et al., 2018). This is due partly to the improved techniques in preparation and high-magnification (400−1000×) microscopy ( Sidorchuk, 2013), and the discovery of major new amber deposits. Taxa described more than a century ago in Eocene Baltic amber are being redescribed in great detail (e.g., Norton, 2000; Sidorchuk and Norton, 2010, 2011), which establish a new standard for study.

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.