
Entomocorus melaphareus, Akama & Ferraris & Jr, 2003
|
publication ID |
https://doi.org/10.1590/S1679-62252003000200001 |
|
DOI |
https://doi.org/10.5281/zenodo.16877262 |
|
persistent identifier |
https://treatment.plazi.org/id/03A187BA-FF8F-2661-8AC9-6744FEDAFD9D |
|
treatment provided by |
Carolina |
|
scientific name |
Entomocorus melaphareus |
| status |
sp. nov. |
Entomocorus melaphareus View in CoL , new species
Holotype. MZUSP 76413 View Materials , 58.6 mm SL, Brazil, Pará, Monte Alegre, Costa do Curuá, left bank of Amazon River , 2 o 13’46.2”S 54 o 01’27.6”W. 7 Mar 2002. A. Akama, C. Lopes & S. Fonteles Santos. GoogleMaps
Paratype. MZUSP 76445 View Materials , 43.1 mm SL, Brazil, Amazonas, between Parintins and Itacoatiara, Amazon River along left bank, at mouth of igarapé, 2 o 31’48.0”S 56 o 58’53.2”W. 13 Mar 2002. A. Akama, C. Lopes & S. Fonteles Santos GoogleMaps .
Diagnosis. A species of Entomocorus having pelvic and pectoral fins densely covered with dark pigment, at least along anterior rays, in contrast to the hyaline fins of its congeners.
Description. [Fin ray counts: D II,5; P I,8; V i,5; A iv,15 ( paratype) or iv,17 ( holotype); C i,7/8,i] Lateral, dorsal, and ventral views in Fig. 1 View Fig illustrate head and body shape, form and position of fins, and pigment pattern. Descriptive morphometric and meristic data presented in Table 1.
Body robust, short; body depth at dorsal-fin origin 24.1- 25.9 % of SL, at anal-fin origin 24.5-24.6 % of SL; body compressed, width at anal-fin origin 32.2 -38.4 % body depth at that point. Lateral line complete and midlateral; canal forms irregular zig-zag pattern, with oblique, short, posteriorly directed branches, but canal does not bifurcate at caudal-fin base.
Head depressed anteriorly; depth at dorsal-fin origin slightly longer than width at cleithrum; profile straight from snout to dorsal-fin origin. Snout length slightly longer than orbital diameter, snout margin rounded. Interorbital width slightly less than one-half of HL (44.4-45.5 %) and almost equal to eye diameter (37.8-38.4 % of HL). Eye large, lateral, visible in both dorsal and ventral views. Barbels slender; maxillary barbel extending past anal-fin origin. Mandibular barbels originate in transverse row at lower jaw margin just lateral to jaw symphysis; inner mental barbel extends to pectoral-fin base; outer mental barbel reaches nearly to middle of pectoral fin. Branchiostegal membrane broadly attached to symphysis. Gill opening relatively wide, ventral margin of opening extends ventral to horizontal through pectoral-spine base. Mouth nearly terminal, upper jaw extends slightly beyond lower jaw. Teeth of upper jaw visible when mouth closed. Jaw teeth minute, barely extending through epidermal tissue; teeth in one row on upper jaw, with additional 1 or 2 rows laterally; tooth band on lower jaw with one row laterally and an irregular tooth patch at symphysis.
Dorsal-fin origin located posterior to vertical through opercular flap. Dorsal-fin base narrow, its length one-third of HL. Dorsal-fin spine slender, straight; anterior margin of spine smooth, posterior margin with medial row of widely spaced and reduced dentations. Dorsal fin with five branched rays; rays slender. Adipose dorsal fin relatively large and triangular.
Caudal fin deeply forked, lobes pointed. Outermost branched rays twice as long as middle rays. Lobes almost symmetrical, upper lobe slightly longer than lower lobe.
Anal-fin base short, its length approximately two times in HL; anal-fin origin located near to vertical through adipose-fin origin. Last unbranched and first branched analfin rays longest; rays decreasing slighthy in length posteriorly. Anal-fin margin straight in paratype, but convex in nuptial males ( holotype; see Sexual dimorphism, below). Anal fin with 4 or 5 unbranched, and 15 to17 branched, rays.
Pelvic fin acutely pointed, first branched ray longest. Pelvic-fin with 1 unbranched, and 5 branched, rays. Pelvic-fin origin located at approximately middle of SL.
Pectoral-fin with 1 stout spine and 8 branched rays. Spine serrated along both margins, antrorse serrae with 16-17 dentations on anterior margin and retrorse serrae with 13-15 dentations on posterior margin. Anterior pectoral-fin rays longest, fin margin straight laterally and convex along medial most rays.
Pigmentation pattern in alcohol. Ground coloration of head and body white. Middorsal line with wide black stripe extending from dorsal-fin origin to base of hypural plate. Lateral and ventral surfaces of body with fine scattered dark pigmentation of variable intensity but more concentrated on dorsal portion of body. Dorsal surface of head with large middorsal black spot extending from posterior limit of cranial fontanel to base of supraoccipital spine. Dorsal surface of snout with dark ovoid spot extending between anterior and posterior nares. Dorsal and lateral surfaces of head with scattered dark chromatophores, especially postorbitally. Dorsal fin with dark pigmentation along interradial membrane between spine and first branched ray and along distal part of anterior one-half of fin margin; remainder of fin pale. Adipose fin with some scattered pigmentation along base. Caudal fin with indistinct band of dark pigmentation at base, pigment somewhat more concentrated dorsally and with diffuse concentration of chromatophores on distal portion of dorsal lobe of fin. Anal fin with few scattered chromatophores at base, extending onto fin anteriorly. Pelvic fin with concentrated dark pigment on most of interradial membranes. Unbranched fin ray with little pigmentation, appearing pale in contrast to remainder of fin. Pectoral fin with dark pigmentation on all but posterior most portion of fin; pigmentation somewhat more concentrated basally. Barbels with scattered dark pigmentation, appearing pale or dusky.
Color in live specimens. The pigmentation pattern described above was observed in the living specimens, however, when alive, the specimens exhibited yellow pigmentation on the dorsal and adipose fins and the dorsal lobe of the caudal fin.
Sexual dimorphism. The holotype of Entomocorus melaphareus is presumed to be a nuptial male, based on its similarity in appearance to nuptial males of Entomocorus gameroi (called terminal dimorphic male by Mago-Leccia, 1983, pers. obs.). The paratype is a juvenile or a nonbreeding male in which the maxillary barbel is ossified, but not fully developed. Differences between the two specimens are presumed to be sexually dimorphic characters, inasmuch as Mago-Leccia (1983) did not report on any externally evident character that distinguished sexually mature females from non-breeding males or juveniles of either sex. The following description is a summary of characteristics of the nuptial male that are usually not found in non-breeding individuals.
The dorsal-fin spine is proportionally more elongate and slender. The adpressed fin extends to past the origin of the adipose fin, rather than falling far short of the adipose-fin origin. The dorsal-fin spine has few weak serrations on the distal half of its anterior margin, and small rounded unculi. The first ray of the pelvic fin is greatly elongated compared with that of the paratype, extending to level of the caudalpeduncle origin. The ray is expanded into a flattened paddle, with the dorsal surface of the paddle densely covered with small, rounded, spiny, anteriorly directed unculi.
The maxillary barbel is enlarged and rigid for its basal one-half, from the expanded ossification of the maxilla. The ossified portion of the barbel is arched and fits into a shallow groove that extends across the ventral margin of the orbit. The dorsal and medial surfaces of the ossified portion of the barbel are covered with unculi. A row of overlaping, scalelike unculi extends from the distal extent of the ossified barbel to near the base, a character previously mentioned by Ferraris & Vari ( 1999). The unculi are small and rounded basally as well as along the medial margin of the barbel.
The snout appears to be broader and somewhat depressed; its appearance apparently due to the enlarged size of the maxilla at the anterolateral margin of the snout. The dorsal surface of the snout is covered with small, rounded unculi, which are larger and denser anteriorly, and extend posteriorly to the nuchal plates.
The anterior serrations of the pectoral-fin spine are curved ventrally, extending well below the horizontal plane of the spine and its posterior serrations. In the non-breeding specimen, all serrations extend anteriorly or posteriorly from the shaft of the spine, but are not curved ventrally. The pectoral-fin spine has clumps of small, rounded unculi on its ventral surface but no unculi were found on the dorsal surface of the spine or on other parts of the pectoral-fin.
The anal-fin is highly modified when compared with the fin of the non-breeding specimen. The base of the three unbranched rays and the three anterior most branched rays are elongated and anteroposteriorly expanded. The distal parts of the expanded rays are either in contact with the more anterior or posterior rays, or nearly so, such that the distal part of the anterior extent of the fin is a rigid, blunty rounded paddle-like structure. The anterior unbranched anal-fin rays that do not contribute to the paddle-like structure are enlarged, but not noticeably elongated. The urogenital opening is at the tip of an elongated, flexible urogenital papilla that originates just anteriorly of the anal-fin origin.
Dermal unculi on this species have a pattern unique among auchenipterids. The presence of dermal unculi on the pectoral, pelvic and dorsal-fin rays is recorded for the first time in auchenipterids. Dermal unculi are also present on the dorsal surface of the cranium, a characteristic of several auchenipterid genera, including Auchenipterus, Parauchenipterus, Epapterus, and Pseudepapterus, although not previously described in any detail. A unique characteristic of Entomocorus is the presence of abdominal unculi, distributed in a “V” like pattern and extends from posterior of isthmus and divides into two lateral branches that flank the abdominal region laterally. We first observed this in the holotype of E. melaphareus and, although Mago-Leccia ( 1983) did not mention the presence of these unculi in his specimens of Entomocorus gameroi, we confirm their presence in the nuptial males examined during this study. It is worth mention that no nuptial males of E. bejamini were found in the examined collections during our comparative study.
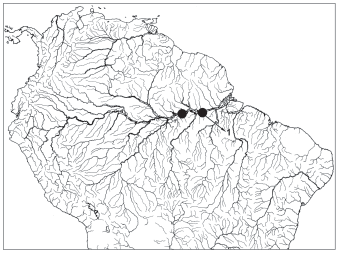
Distribution. Known only from two localities on the left bank of the Amazon River below Manaus ( Fig. 3 View Fig 3 ).
Remarks. This species is quite similar in appearance to its two congeners, Entomocorus benjamini and E. gameroi . Meristically, the three species are not distinguishable and are most readily diagnosed on color pattern differences. There were no obvious morphometric differences that could be demonstrated with the limited sample size of the new species.
The new species is more similar in coloration to Entomocorus benjamini than it is to E. gameroi . Entomocorus melaphaereus and E. benjamini share similar marking on the caudal fin, in which the upper lobe has an indistinct blotch of dark pigment and the caudal-fin base has a diffuse vertical bar. In contrast, E. gameroi has a broad, nearly horizontal stripe of dark pigmentation that extends from the base to the tip of the upper lobe of the caudal fin. In addition, the lateral surfaces of the body of E. melaphaereus and E. benjamini has little, if any dark pigmentation, whereas E. gameroi sometimes has irregular blotches of dark pigmentation and, often a dark midlateral stripe. However, despite the similarity in appearance of E. melaphaereus and E. benjamini , even darkly pigmented specimens of E. benjamini lack the black pigmentation on the pectoral and pelvic fins that characterize the new species.
Etymology. The name melaphareus is derived from the Greek melanos, black, and aphareus, pelvic fin of a tuna, in reference to the unique condition in this species, in which the pelvic fin is dark.
Comparative material. Entomocorus gameroi . Venezuela: Apure: USNM 258187 About USNM , 1, 47 mm SL, not transformed, side channel of río Apure , ca. 5 km west of San Fernando de Apure, 7°53’N 67°29’W GoogleMaps ; USNM 258189 About USNM , 1, 38 mm SL, not transformed, caño 1 km south of ferry crossing on río Apurito , where crossed by road from San Fernando to Cunaviche, 7°33’N 67°38’W GoogleMaps ; USNM 258229 About USNM , 11, 33-45 mm SL, none transformed, río El Cañito where crossed by road from San Fernando to Cunaviche, 7°28’N 67°39’W; GoogleMaps Anzoategui: ANSP 166520 About ANSP , 3 / 36, 57-59 mm SL, transforming males, río Orinoco basin, Soledad, L.Terecaya, 8°11’30"N 63°27’20"W; GoogleMaps Bolivar: ANSP 178475 About ANSP , 1, fully transformed male, río Cunaviche , ca. 20 km SW of Cunaviche on San Fernando de Apure to Puerto Paez Hwy , 7°20’00"N 67°35’00"W; GoogleMaps Guarico: ANSP 158849 About ANSP , 2, 51-58 mm SL, fully transformed males, Presiamos lagoons between La Antena y caño Falcon. GoogleMaps USNM 258192 About USNM , 3, 38-41 mm SL, none transformed, caño Falcon, río Portuguesa drainage basin, 8°14’N 67°35’W GoogleMaps ; USNM 258225 About USNM , 7, 38-43 mm SL, none tranformed, caño to west of highway from Calobozo to San Fernando, about 35 km south of Fundo Masaguaral (caño Falcon), 8°14’N 67°35’W GoogleMaps ; Portuguesa: USNM 348676 About USNM , 19, 32- 44 mm SL, none transformed, Guanare-Guanarito road at road km 60, 8°49’39"N 69°20’42"W GoogleMaps . Entomocorus benjamini . Bolivia: Dept. Beni, Ballivia Prov., USNM 305608 About USNM , 3, 45-53 mm SL, none transformed, río Curiraba , 10 km NE El Porvenir Biol. Sta. , 40 Air km E San Borja, 14°05’S 66°17’W GoogleMaps . Entomocorus sp. Brazil: Mato Grosso do Sul: MZUSP 59465 View Materials , 24, 32-46 mm SL, none transformed, Corumbá, left bank of rio Vermelho , 19º37’22”S 56º57’27”W GoogleMaps ; MZUSP 59649 View Materials , 7, 30-52 mm SL, none transformed, Aquidauana, rio Taboco (swamp), 20º4’12”S 55º38’37”W GoogleMaps .


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
ParvPhylum |
Osteichthyes |
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |