
Cataglyphis hannae, Agosti, 1994
|
publication ID |
https://doi.org/ 10.5281/zenodo.2949937 |
|
DOI |
https://doi.org/10.5281/zenodo.5681917 |
|
persistent identifier |
https://treatment.plazi.org/id/03A1879C-FFD3-FFB4-CD75-987266B6FD85 |
|
treatment provided by |
Plazi |
|
scientific name |
Cataglyphis hannae |
| status |
sp. nov. |
Cataglyphis hannae View in CoL n. sp.
Material examined
Holotype: <?, HL 1-40, HW 1-20, EL 0-50, SL 218, AL 2-63, CI 86, SI 181, El 41-7, FI 35-4, LI 100-8, TAI 114-3.
Diagnosis of male
Ant of the Cataglyphis bicolor species group ( Agosti, 1990), with the following diagnostic features:
(1) Black head and alitrunk.
(2) Alitrunk length 2-63 mm.
(3) Subgenital plate distally trilobed with the median process triangular and not digitiform.
(4) In ventral view, median process with hair-carrying pits to the tip.
(5) In lateral view, apicolateral appendix of sagitta not overreaching outline of apical, serrated plate.
(6) Apicolateral appendix of sagitta short and terminally rounded; in ventral view only slightly raised above the plane of the apical, serrated plate.
(7) Long hind tibiae (TAI 114-3).
Paratype: ♂ ♂ HL 1-30-1-43, HW 1-05-1-19, EL 0-44-0-50, SL 1-98-2-23, AL 2-48-2-73, CI 80-8-86-6, SI 175-5-195-6, El 41-1-43-0, FI 91-4-103-2, LI 101-7-105-5, TAI 108-3-116-0 (n = 8).
There is a slight variation in size, but the extremes of the range of absolute measurements are always far below those of the other males of the bicolor species group. Compared with all the other males of the bicolor group, hannae has the longest hind tibiae (TAI> 108). Head and alitrunk always black in colour.
Diagnosis offemale
Ant of the Cataglyphis bicolor species group with the following diagnostic features ( Fig. 1B View Fig ):
(1) Small size, alitrunk <3-2 mm.
(2) Scape much longer than head length (SI> 120).
(3) Long hind tibiae (TAI> 90).
(4) Head and alitrunk bright red.
(5) First funicular segment of intermediate length (28 <FI <35).
(6) Low rounded petiole in lateral view.
Paratypes: ♀ ♀ HL 1-75-1-88, HW 1-65-1 -73, EL 0-48-0-50, SL 2-00-2-18, AL 2.95-3.13, CI 90-4—94-3, SI 121-2-129-9, FI 29-0-31-1, El 28-4-29-0, LI 101-8-107-1, TAI 93-5-100-8 (n = 5).
Deposition of type material
Holotype: ♂, Tunisia, El Guettar, 34.33°N, 8.92°E, 300 m; in Oasis, at the edge of irrigated fields under Eucalyptus trees, D. Agosti, 0 8.06.1992, sample F92039 View Materials , in nest with Cataglyphis bicolor ( MHNG). GoogleMaps
Paratypes: 5 ♀ ♀, (alates) and 14 ♂ ♂, same as holotype. 7 males, Tunisia, El Guettar, 34.33°N, 8.92°E, 300 m; in Oasis, at the edge of an irrigated lucerne field, D. Agosti, 0 8.06.1992, sample F92309 View Materials , in nest with Cataglyphis bicolor ( BMNH, CDA, CRW, MCZ, MHNG). GoogleMaps
C. hannae can be separated from all the other known species in the bicolor group by the above mentioned diagnostic characters, especially by the body size, the long antennal scape and hind tibiae, the low rounded petiole, and the male genitalia. C. abyssiniens , the only species with small females, has a much shorter scape (SI <113), and shorter hind tibiae (TAI <85); no males of this species are known. The long black hairs on the occiput of the head as seen in bicolor are not present in hannae , and thus exclude the possibility of hannae being a microgyne of bicolor .
Although> 1500 different samples of species of the bicolor complex are in our collections, and most of them from Tunisia, no further hannae specimens have been collected before. The host of hannae is distributed along the southern foothills of the Atlas mountains in the transitional zone from the Mediterranean to the Desert region, not reaching the Atlantic in the West and not extending into Libya (Agosti et al., in preparation). Thus, hannae might be more common, certainly if one considers that the southern extension of the Mediterranean vegetation during the Pleistocene reached as far south as the Saharan mountains Tassili and Hoggar ( Quezel, 1965). But social parasitic ants tend to have a clumped distribution ( Buschinger, 1985) and the restricted collecting area might truly reflect its distribution. Finally, these males and females were collected as alates, during a survey where> 200 nests were at least partially dug up.
The habitat was at the edge of an irrigated Medicago sativa field and on a sandy place in the shade of some tall Eucalyptus trees on the side of a track within the oasis. In this desert region, the occurrence of species of the bicolor group is restricted to oases.
The position of hannae within the bicolor species group is established by the male genitalia and the funicular index of the female. More problematic, and challenging, is the sister group relationship within the bicolor group. A phylogenetic analysis using morphological characters of nodus (outgroup), abyssinicus, bicolor, savignyi, viaticus ( Table 1 View Table 1 ) and the explicit enumeration procedure from the computer program Hennig86 ( Farris, 1988), resulted in 3 most parsimonious trees. Subsequent successive weighting established 2 most parsimonious trees ( Fig. 2 View Fig ). Both these trees share the sister group hannae + abyssinicus, and support a sister group viaticus + (hannae + abyssinicus). The sister group relationship of hannae + abyssinicus is so far only supported by female characters, as the males of abyssinicus are unknown. However, the long antennal scape and the small size of the females of abyssinicus , which is the same as in hannae and unique in species of the bicolor species group, indicate an inquiline life style for this species too. A more detailed discussion will be provided by Agosti (in preparation).
As all the results are based on morphological characters, it would be interesting to test them using other characters such as DNA sequences, electrophoretic data or the chemical composition of glandular compounds, all of which have already showed their specificity at the species level ( Keegans et al., 1992; Agosti et al., in preparation).

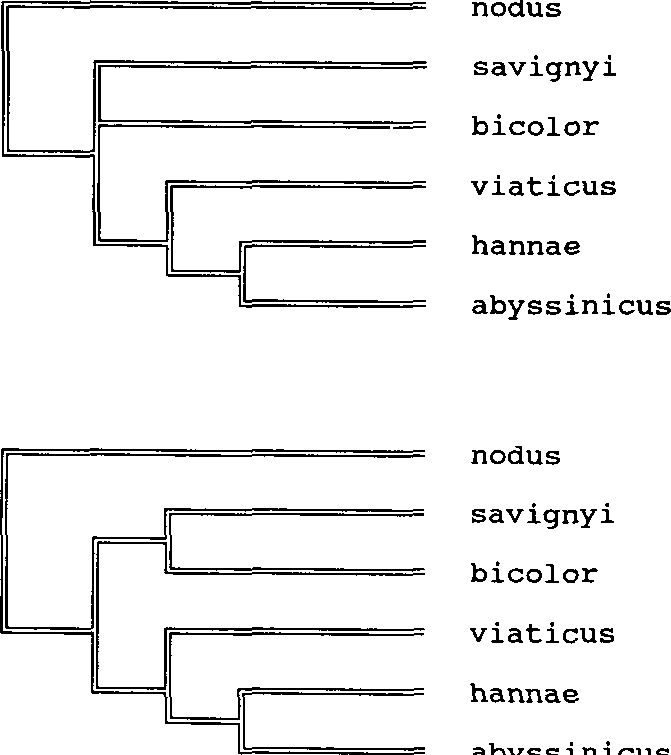
Table 1. Data matrix. One ri: SI> 170 (character state 0), SI <170 (1); 2 Queen: CI> 95 (0), C K 95 (1); 3 Q: SI<105 (0); SI>105 (1); 4 Q, TL<3-5mm (0), TL<3-5mm (1); 5 Q, TAI <90 (0), TAI> 90 (1); 6, Q, white hairs on occiput of head (0), black hairs (1); 7, Q, head and alitrunk bright red (0), dark red to black (1); 8 M, lateral apical appendage of subgenital plate digitiforme, parallel sided (0), triangular (1); 9 M, median apical appendage of subgenital plate in crossection (at midlength) rounded (0), angular (1); 10 M, Sagitta with antero-lateral appendage convergent at base (0), parallel sided at base (1); 11 M, in lateral view, antero-lateral appendage of sagitta crossing outline of apical blade (0), not extended beyond (1); 12 M, median appendage of subgenital plate shorter as lateral appendages (0), of same length (1); 13 M, antero-lateral appendage of sagitta apically pointed (0), rounded (1). (A full discussion of the characters, and coding, based on> 1000 specimens, will be provided elsewhere (Agosti, in preparation).
| Species | Characters | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | |
| Nodus | 1 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 1 | 1 |
| Hannae | 0 | 1 | 1 | 1 | 1 | 0 | 1 | 1 | 0 | 0 | 1 | 0 | 0 |
| Abyssinicus | - | 1 | 1 | 1 | 0 | 0 | 1 | - | - | - | - | - | - |
| Bicolor | 1 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 1 | 1 | 0 | 0 | 0 |
| Savignyi | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 |
| Viaticus | 1 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 |
| MHNG |
Switzerland, Geneva, Museum d'Histoire Naturelle |
| BMNH |
United Kingdom, London, The Natural History Museum [formerly British Museum (Natural History)] |
| CRW |
CRW |
| MCZ |
USA, Massachusetts, Cambridge, Harvard University, Museum of Comparative Zoology |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |