
Henricia leviuscula ( Stimpson 1857 )
|
publication ID |
https://doi.org/ 10.5281/zenodo.275445 |
|
DOI |
https://doi.org/10.5281/zenodo.6205506 |
|
persistent identifier |
https://treatment.plazi.org/id/03A0E238-AE76-FFF7-FF42-023BFBC2294C |
|
treatment provided by |
Plazi |
|
scientific name |
Henricia leviuscula ( Stimpson 1857 ) |
| status |
|
Henricia leviuscula ( Stimpson 1857) View in CoL
Figs. 2 View FIGURE 2 A–E, 3A, 4A, 5A, 5C, 5E
Type: USNM 0 3357 (holotype)
Type Locality: Puget Sound, Washington, USA
Linckia leviuscula Stimpson, 1857: 529 .
? Cribrella leviuscula (Stimpson) , Cribrella laeviuscula (Stimpson) , or Henricia leviuscula (Stimpson) View in CoL : Numerous authors reviewed by Fisher, 1911 and by A. Clark, 1996.
Henricia leviuscula ( Stimpson 1857) View in CoL : in part Fisher, 1910: 570; in part Verrill, 1914: 215; in part Djakonov 1961:23, Pl. II, fig 9; in part Lambert, 1981: 100 –103; 2000: 104–106.
Henricia leviuscula View in CoL “var. A after Fisher”: Djakonov 1950: 95, f. 79, 80
Henricia leviuscula View in CoL var. B: in part Fisher 1911: 280 –291, pl. 70, f. 2, pl. 111, f. 6
Henricia leviuscula View in CoL var. leviuscula : in part Verrill 1914: 217, pl. XII–f. 5, pl. XIII, f. 1–2.
Henricia leviuscula View in CoL var. lunula : in part Verrill 1914: 219
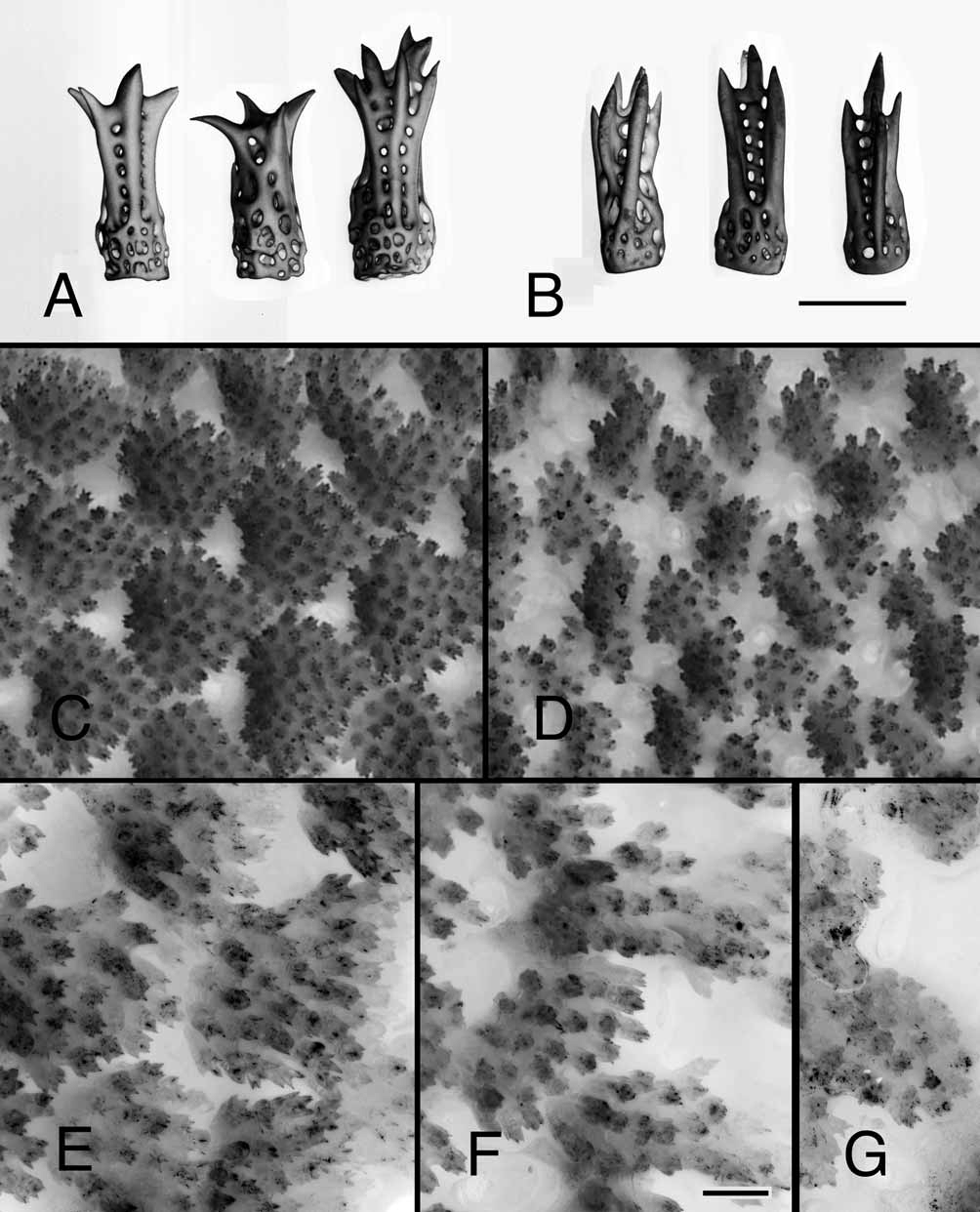
Type material examined: A specimen ( Fig. 2 View FIGURE 2 A–E) originally labeled only “ Henricia leviuscula, Id. W.K. Fisher, Ft. Steilacoom [near Tacoma, Pierce Co., Washington; 47°10.80΄N 122°35.42΄W], Pudget [sic] Sound, Dr. Geo. Suckley Col.” matched Stimpson’s brief published locality: “Found in Puget Sound, by Dr. Suckley.” Our analysis revealed this specimen to be the likely holotype and it has since been labeled as such in the USNM collection.
Other material examined: more than 40 specimens in our collections from the San Juan Archipelago, including USNM 1116587 ( Figs. 3 View FIGURE 3 A, 4A, 5C, 5E), Cattle Pt., San Juan Id., San Juan Co., Washington, intertidal, and MFS 149 ( Fig. 5A View FIGURE 5. A – B ), and two from Lovit Island, British Columbia. Museum specimens identified included: USNM 5991 (Baranoff Id., Alaska, 18m); USNM 3386 and 9225 (Puget Sound); USNM E4679 (“probably Alaska”); USNM E8678 (Departure Bay, British Columbia); USNM E3780 (Mouth of Strait of Juan de Fuca, 73 m); USNM E3820 (Port Townsend, Washington, 27–48 m); CASIZ 0 0 8559 (1 of 9 previously designated as vouchers for Henricia leviuscula by Fisher (1911:281–282, 285)); CASIZ 112955 (Anchor Pt., Kenai Peninsula, Alaska); CASIZ 180558 (San Juan Channel off Lopez Id., San Juan Co., Washington, 100–110); LACM 1959-281.3 (Monterey, California); LACM 1966-36.16 (lot of 2, off Cowichan, Sydney Channel, British Columbia 18–37 m); LACM 1966-37.19 (lot of 2, Sister’s Rocks, outer Squamish Harbor, Hood Canal, Washington, intertidal); LACM 1966.39.23 (lot of 2, MacKaye Harbor, Lopez Id., Washington, intertidal); YPM 9836 (1 of 2 syntypes for Henricia leviuscula (Stimpson) var. lunula Verrill, 1914, Victoria, British Columbia, Canada, 20m).
Description: Most specimens have 5 slender rays, but 6-rayed individuals occur. Slender rays are without basal swelling and taper evenly to the ray tip. The dried holotype has 5 slender rays, not basally swollen, with R about 35 mm and r about 6.3 mm, R/r = 5.6 ( Fig. 2 View FIGURE 2 A–E). Of 42 specimens from the San Juan Archipelago and 2 from Lovit Id., British Columbia, the largest had R = 70 mm, r = 10 mm and the smallest, R = 22 mm, r = 4.0 mm. The range of R/r was 5.0 to 8.3.
The aboral color in living specimens is distinctive bright orange, sometimes slightly red-orange ( Fig. 3 View FIGURE 3 A), with the color characteristically continuing onto the oral surface, fading to light orange or yellow sometimes over the ventrolateral plates but most usually on only the adambulacral plates.
The aboral surface appears smoothly and regularly paved or scaled, this surface being composed of closeset pseudopaxillae with narrow intervening spaces and small recessed papular areas. In outline the pseudopaxillae are oblong or irregularly polygonal on the disc and oval, crescentic, heart-shaped, or triangular on the rays, with the concave or flat side toward the disc ( Fig. 2 View FIGURE 2 B, 2D, 4A, 5C, 5E). The madreporite is not particularly prominent and bears single rows of spines that are like those on the pseudopaxillae.
Aboral papular areas have 1, sometimes 2, rarely 3 papulae. A regular series of single papulae occurs on the oral surface between the inferomarginal and ventrolateral plates on the disc and most of the ray length. A few single papulae may occur in an irregular series between some ventrolateral and adambulacral plates.
Aboral pseudopaxillae are composed of spines set in tissue-covered, close-packed, upright arrays of 20 to 100, but usually 25 to 55 ( Fig. 5 View FIGURE 5. A – B C, 5E). In living specimens the pseudopaxillae are covered by pigmented tissue through which some of the radiating glassy thorny tips of the spines might just be discerned. The aboral spines are stout with fenestrated glassy shafts, commonly 80 to 100 µm in diameter, with smooth sides. The ends of the spines have 3 to 7 heavy thorn-like points or tips (as noted by Djakonov 1961), of about equal sizes that flare in a fairly even radiating pattern that spans 100 to 190 µm ( Figs. 2 View FIGURE 2 D, 5A, 5C, 5E). A large spine on the holotype is about 220 µm long, has a shaft about 70 µm in diameter, and ends in prominent sharp points radiating over a span up to 165 µm.
Ray sides have the lowermost dorsolateral pseudopaxillae in approximate 1:1 alignment with the superomarginal pseudopaxillae (plates) below them, but in offset alignment with the pseudopaxillae above them. Superomarginal pseudopaxillae are larger than the dorsolaterals, sometimes as much as twice the size.
Ray lower sides have three conspicuous series of pseudopaxillae: rectangular superomarginals and inferomarginals and smaller, squarish ventrolaterals. These series of pseudopaxillae are aligned in 1:1 correspondence beside the adambulacral plates for most of the ray length. The superomarginal series descends abruptly in the interradial area to lie adjacent to the inferomarginal series, enclosing a very small interradial triangle of only a few intermediate plates and, as Hayashi (1940) noted, there are no secondary ventrolaterals, singly or in series. The ventrolateral series usually extends most of the ray length, but sometimes only half. Among 21 specimens, the ratio of ventrolateral to adambulacral plates (V/A, counted from mouth to ray tip) ranged from 0.43 to 0.85 but only 2 specimens had ratios of 0.5 or lower.
Near the base of the ray, a superomarginal pseudopaxilla bears 25 to 100 spines; an inferomarginal, 32 to 100 spines; and a ventrolateral, 5 to 20 spines. The spine count varies among individuals and is not correlated with body size. Each adambulacral plate ( Fig. 2 View FIGURE 2 C, 2E) bears 1 thin curved deep-furrow spine and 8 to 17 large, slightly curved, columnar spines in a zigzag or double row that becomes a triple or quadruple row on the edge farthest from the ambulacral furrow. The larger spines near the furrow are blunt-ended with finely spinous surfaces; those farther from the furrow are smaller and more coarsely spinous with sharp terminal points but not radiating thorns. On the holotype, the largest adambulacral spine is about 175 µm in diameter and 775 µm long.
Distribution: H. leviuscula , as restricted herein, has a confirmed range limited to southern Alaska (Kenai Pt.) south to Puget Sound (see Remarks). In the San Juan Archipelago it is uncommon in rocky intertidal areas but occurs more often in shallow subtidal areas. Many of the museum specimens whose identity we have confirmed are from the San Juan Archipelago, northern Puget Sound, or Vancouver Island.
Reproduction. We assume that sexes are separate. When specimens of H. leviuscula , or one of a number of similar co-occurring species, have been spawned in isolation, eggs have not been observed to develop. Spawned eggs have been measured at 1342 µm diameter ( Strathmann et al., 2002), and we confirmed that one female induced to spawn released slightly ovoid red-brown eggs 1.45 x 1.16 mm in diameter. Spawning individuals have been seen with the disc elevated to shed sperm or eggs freely into the water through aboral gonopores ( Strathmann, 1987). Spawning season, like that of congeners with pelagic development, is probably early spring in the San Juan Archipelago. Development is similar to that described for other Henricia species ( Masterman 1902, Strathmann 1987, Komatsu et al. 1995). Embryonic stages do not adhere to one another but float freely. Post-hatching larvae are ciliated and swim. Future reproductive observations need to be considered with respect to knowledge of multiple H. leviuscula -like species, but no one has ever reported feeding in any Henricia larva. This and the large size of their yolky eggs implies that larvae of H. leviuscula are lecithotrophic. However, the possibility that Henricia larvae might uptake dissolved organic matter has not been tested and no one has sectioned Henricia larvae to see if some might have an open mouth. The brachiolar larva has an anterior lobe with three brachiolar arms surrounding an adhesive disc and late in development has five groups of podia protruding on the left side. Metamorphosis begins after about 30 to 40 days; the anterior lobe is resorbed into the oral edge of the disc.
Remarks. We discovered a dried specimen in the USNM collections that matches the published record and we consider it to be the holotype. This specimen may be the one Fisher (1911: 284) wrote of as “perhaps the type; if not, it is at least a topotype.” In describing a multi-species collection from Puget Sound made by N. R. Harrington from Columbia University in 1896 and 1897, H. L. Clark (1901) introduced two nominal varieties for Stimpson’s leviuscula (as “ laeviuscula ”): crassa and attenuata . He provided only a brief diagnosis for each involving a ratio of the distance from the disc center to either the ray tip (R) or the disc edge between rays (r). R/r ranged from 2.0 to 3.5, n = 25 for crassa vs. R/r = 5.0, n = 1 for attenuata . We have been unable to locate Clark’s type material at either the American Museum of Natural History (affiliated with Columbia University) or MCZ (where Clark later worked). Given the insufficient nature of Clark’s characterization and the absence of known type material, we consider these nominal varieties as nomen dubia . Verrill’s (1914) description of the color of H. leviuscula var. attenuata ( Clark 1901) as “orange and orange-red above, yellow below” does not apply to H. leviuscula as restricted herein.
We consider Fisher's H. leviuscula variety B to correspond in part to H. leviuscula based on his statement (1911:281) that variety B is typical of the “littoral form from Puget Sound with very regular marginal plates, eight to fifteen adambulacral spinelets, and the abactinal spinelets ending in numerous sharp denticulations rather than in a solid glassy tip.” He had examined “a specimen evidently labeled by Stimpson,” and wrote that “the type is therefore one of the various forms of variety B, among those listed herewith.” His photographs ( Fisher, 1911: pl. 70, fig. b, c) are not at high enough magnification to show the form of the “sharp denticulations” of the aboral spines. Nothing in his description of variety B is incompatible with H. leviuscula , but his variety B may include at least one other sympatric species that has subtly different spines. We re-examined many museum specimens identified as H. leviuscula by Fisher and found that most lack the characteristic aboral spines exhibited by the holotype and belong instead to other species, some of which are likely undescribed. We believe that most reports of H. leviuscula from the North Pacific refer to other similar species. Fisher's H. leviuscula varieties A and C to F are probably all other species, some of which will be assessed in a regional revision of the genus (D. Eernisse and M. Strathmann, in prep.).
Verrill's (1914) eight varieties and one subspecies of leviuscula are mostly other species. We were able examine the only two known syntypes of H. leviuscula ( Stimpson, 1857) var. lunula Verrill, 1914 and have tentatively identified one of these (YPM 9836), from Victoria, Vancouver Island, as H. leviuscula . We designate YPM 9836 as a paralectotype of H. leviuscula . We designate the other syntype (YPM 9835) as lectotype of lunula because this is clearly the only specimen figured and the only lunula “ type ” mentioned by Verrill. The aboral plates denuded of spines accord with his figures 2a–b (Plate 88) and the specimen is from Monterey, California, as stated by Verrill in reference to the figured specimen (p. 218); however, it is unclear whether he considered both specimens in the lot as “ types ” or only the figured specimen as holotype in a modern sense. Given the uncertainty, a lectotype designation is appropriate but the lectotype is not H. leviuscula and will be treated in a subsequent study (D. Eernisse and M. Strathmann, in prep.).
Djakonov (1950: 95) distinguished two varieties of H. leviuscula based on form of aboral spines. The aboral spines he described for H. leviuscula variety A, as “low with sharp but coarse glassy spines protruding on various sides as found in a specimen from Nanaimo Bay” ( Djakonov 1950: figs. 79, 80), corresponds to the spine morphology we found on the rediscovered holotype. We have not seen any specimens studied by Djakonov but his H. leviuscula variety A could be H. leviuscula . It should be noted, however, that Djakonov's variety A is not equivalent to Fisher's variety A, which had aboral spines ending in a solid glassy tip. Djakonov's H. leviuscula variety B, characterized by aboral spines “stout with a thorny apical knob as found on typical specimens from Monterey Bay” ( Djakonov, 1950: figs. 83, 84) is not H. leviuscula but another species possibly still undescribed.
Fisher (1911) reported that leviuscula in “typical form” (his variety B) occurred from the Aleutian Islands, Alaska, to Monterey Bay, California, and that southern specimens that were not “typical” extended to Santa Barbara Channel and San Diego, California. In contrast, Verrill (1914) considered H. leviuscula to be a “southern” form, noting that most of Fisher's specimens were from sites south of Sitka, Alaska. We have confirmed Fisher's identification of some H. leviuscula from near Sitka and Fort Wrangel (Alaska), Port Renfrew (Vancouver Island, British Columbia), Port Townsend (North Puget Sound), and Hecata (= Heceta) Bank off Oregon. Djakonov (1950, figs. 79, 80) described and illustrated his variety A from a specimen from Nanaimo, British Columbia and said that it had spines like those of the holotype.
Uchida (1928) described a single specimen of leviuscula from Japan, but Hayashi (1940) compared material from Japan with specimens thought to be of leviuscula from Kodiak, Alaska, and concluded that leviuscula was not present in collections from Japanese waters. Djakonov (1950) reported leviuscula from the Sea of Japan, but later ( Djakonov 1961: 23), having decided that at least two species were known under the name, stated it difficult to define the true range, which might include Sea of Japan and near Paramushir Island of the Kurile Islands. Hayashi (1973: 11) did not list leviuscula among seastars from Japan. We did not find leviuscula in a small sample of representative species from Japan kindly made available to us by Mr. Masaki Saba and Mr. Yasuo Kano through the courtesy of Dr. Chitaro Oguro and Dr. Mieko Komatsu.
H. leviuscula View in CoL is distinct from congeners described from Japanese waters ( Hayashi 1940, 1973) or the North Atlantic ( Heding 1935; Rasmussen1965; Madsen 1987, A. M. Clark & Downey 1992), and we have not seen any specimens of H. leviuscula View in CoL as restricted herein from north or west of Kenai Peninsula, Alaska or south of Puget Sound, Washington. In the northern part of its range, in southeastern Alaska, it appears to be less common than its similar congeners (D. Eernisse and M. Strathmann, in prep.) and it is likely endemic to northeastern Pacific cool temperate coastal areas (northern Oregonian province).


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Henricia leviuscula ( Stimpson 1857 )
| Eernisse, Douglas J., Strathmann, Megumi F. & Strathmann, Richard R. 2010 |
Henricia leviuscula
| Djakonov 1950: 95 |
Henricia leviuscula
| Verrill 1914: 217 |
Henricia leviuscula
| Verrill 1914: 219 |
Henricia leviuscula
| Fisher 1911: 280 |
Henricia leviuscula (
| Lambert 1981: 100 |
| Djakonov 1961: 23 |
| Verrill 1914: 215 |
| Fisher 1910: 570 |
Linckia leviuscula
| Stimpson 1857: 529 |