
Neoromicia stanleyi, Goodman, Steven M., Kearney, Teresa, Ratsimbazafy, Malalatiana Michèle & Hassanin, Alexandre, 2017
|
publication ID |
https://doi.org/10.11646/zootaxa.4236.2.10 |
|
publication LSID |
lsid:zoobank.org:pub:DA87FB16-A7E7-43CB-8B3A-0AF1497B231F |
|
DOI |
https://doi.org/10.5281/zenodo.3860418 |
|
persistent identifier |
https://treatment.plazi.org/id/03A087F2-FFAB-FFDF-FF2A-001ED4A20DC0 |
|
treatment provided by |
Plazi |
|
scientific name |
Neoromicia stanleyi |
| status |
sp. nov. |
Neoromicia stanleyi View in CoL sp. nov.
Synonyms
Eptesicus melckorum Hayman & Hill, 1971 View in CoL , in part Eptesicus ( Neoromicia) melckorum Koopman, 1993 View in CoL , in part Neoromicia melckorum Simmons, 2005 View in CoL , in part Neoromicia cf. melckorum Monadjem et al., 2010 View in CoL Pipistrellus ( Neoromicia) melckorum Hill & Harrison, 1987 , in part Pipistrellus cf. melckorum Rautenbach et al., 1993 Pipistrellus cf. melckorum Kearney, 2013a
Holotype. TM 48487, adult male prepared as a fluid preserved specimen with skull removed and cleaned, as well as the baculum removed and cleared. Original field number Teresa Kearney and Ernest C. J. Seamark ( ECJS) 48/ 2009. The holotype was used in the morphological, bacular, and molecular comparisons presented herein. A karyotype is not available for this specimen.
Type locality. Botswana: Okavango Delta, Chitabe, Gomoti River, Highbanks , 19.41979’S, 23.40413°E, 950 m above sea level ( Figure 3 View FIGURE 3 ). The animal was captured on 11 April 2009.
Referred specimens that were sequenced (see Figure 3 View FIGURE 3 for geographical locations): FMNH 97211 About FMNH , adult female, same as type locality; TM 48 484 ( ECJS- 02 /2009), adult female, Botswana: Okavango Delta, Chitabe, Gomoti River , film camp, 9 April 2009, 19.42585°E, 23.41696°S, 950 m above sea level GoogleMaps ; TM 41860, 41861, and 41862, all adult females, Zimbabwe: Mana Pools National Park , all 1 July 1990, 15.7333°S, 29.35°E, 365 m above sea level GoogleMaps ; and NMZL ECJS- 114 /2010, adult male, Zambia: Kafue National Park, Lufupa Camp, confluence Lufupa and Kafue Rivers , 31 December 2010, 14.61694°S, 26.19113°E, 1099 m GoogleMaps . The baculum of NMZL ECJS-114/2010 has been previously illustrated ( Goodman et al. 2015, their Figure 7 View FIGURE 7 G).
Non-sequenced specimens referred to N. stanleyi based on morphology (m) or karyology (k) (see Figure 3 View FIGURE 3 for some of the geographical locations): BMNH 66.546 , unsexed, Zambia: Matebele Plain, 21 July 1964, 16°25’S, 23°08’E (m) GoogleMaps ; BMNH 66.5461 ( W.F.H. Ansell 22944), male, Zambia: between Nyimba and Fort Jameson [= Chipata], 6 August 1964, "found dead on radiator of Land Rover after night journey" (m); TM 36778, male, South Africa : Limpopo, Kruger National Park, Figtree Camp, 26 January 1984, 22°25’50’’S, 31°11’50’’E (k); TM 37844, female, South Africa GoogleMaps : Limpopo, Kruger National Park, Culling Camp, 2 February 1985, 22°25’S, 31°15’E (k); TM 37852, South Africa GoogleMaps : Limpopo, Kruger National Park, Mockford’s Garden, 22°25’S, 31°18’E (k); TM 37858, female, 2 May 1985, South Africa GoogleMaps : Limpopo, Kruger National Park, New Figtree Forest , 22°25’S, 31°18’E (k); TM 37906, male, 4 November 1985, South Africa GoogleMaps : Limpopo, Kruger National Park, New Figtree Forest , 22°25’S, 31°18’E (k); TM 37923, female, 4 December 1985, South Africa GoogleMaps : Limpopo, Kruger National Park, Old Picnic Site, 22°25’S, 31°18’ E (k); TM 37924, male, 4 December 1985, South Africa GoogleMaps : Limpopo, Kruger National Park, Old Picnic Site, 22°25’S, 31°18’E (k); TM 37937, male, 13 April 1985, South Africa GoogleMaps : Limpopo, Kruger National Park, Culling Camp, 22°25’S, 31°15’E (k); TM 37943-37945, all males, 17 April 1985, South Africa GoogleMaps : Limpopo, Kruger National Park , Anthrax Camp, 22°25’S, 31°12’E (k). GoogleMaps
A number of other specimens originally referred to Neoromicia cf. melckorum are presented in Monadjem et al. (2010). Those that have not been listed above include: Botswana ( NMZB 59307), Malawi (in reference to information in Happold et al. 1987), South Africa (TM 34185), Zambia ( NMZB 20019, NMZB 20310, UZMZ 66), and Zimbabwe ( NMZB 30065, NMZB 33756), which we have not been able to examine in the context of this study. Further, Monadjem et al. (2010) illustrate TM 37924 (their Figure 277) and identify the specimen as Neoromicia cf. melckorum , based on karyotype information associated with the specimen; this specimen is not listed under N. cf. melckorum in their appendix of examined specimens. Another ten specimens from the northern part of the Kruger National Park in South Africa (TM 34185, female, TM 34186, female, TM 34240, male, TM 34263, female—South Africa: Limpopo, Kruger National Park, 4 km W Bridge, Levuvhu Hippo Pool, 31°11’E, 22°26’S; TM 37680, female, TM 38599, male—South Africa: Limpopo, Kruger National Park, Figtree Forest, 31°15’E, 22°25’S; TM 37833, female, South Africa: Limpopo, Kruger National Park, Culling Camp, 31°15’E, 22°25’S; TM 38132, female, South Africa: Limpopo, Kruger National Park, Manxeba Windmill, 31°14’E, 22°24’S; TM 38843, female—South Africa: Limpopo, Kruger National Park, New Figtree Forest, 31°18’E, 22°25’S; and TM 39506, male—South Africa: Limpopo, Kruger National Park, Old Picnic Site, 31°18’E, 22°25’S) were identified as N. cf. melckorum based on cranio-dental morphology in Kearney (2005). Thorn (1988) mentioned two specimens from Mozambique ( USNM 365408 and 365409) that have the same dark color to the base of the hair as in BMNH 66.5460 and 66.5461 from Zambia and might be referable to N. stanleyi . Hill and Harrison (1987) illustrated the baculum of BM 83.216 from Mucheni Gorge in Zimbabwe that on its overall size might also be referable to N. stanleyi .
Etymology. We name this species in honor of the late William (“Bill”) T. Stanley, who conducted active field and systematic research on African small mammals, particularly in east Africa, until his sudden death in late 2015 while working in the Ethiopia Highlands ( Newmark 2016). He was the mammal collection manager at the Field Museum of Natural History from 1989 to 2015. Bill was well known amongst colleagues for his vivacious manner and enthusiasm, curatorial skills, and knowledge on African mammals. He readily shared specimens in collaborative projects with numerous colleagues. With his passing, mammalogy has lost a passionate field biologist and museum curator. We propose the English common name “Stanley’s serotine” and in French “sérotine de Stanley”.
Diagnosis. Excluding the genus Scotophilus , Neoromicia stanleyi is a notably large Vespertilioninae and one of the bigger African species within the Pipistrellus - Neoromicia - Hypsugo complex. The dorsal pelage is distinctly long and pale brown to dark brown in coloration. The ventral surface is chocolate brown to silvery-brown on chin and throat, merging to creamy to brownish-cream on chest and belly, and cream or grayish-white towards the venter ( Kearney 2013a). The wings are brown, dark brown or blackish brown, slightly translucent, and with and without white hind border; interfemoral membrane paler and more translucent than wings; and naked skin around muzzle and eyes with a pinkish tinge based on a color photograph ( Figure 4 View FIGURE 4 ). Forearm length in holotype is 36.8 mm and the range in genetically types animals referred to this species is 34–39 (n=8) ( Table 4 View TABLE 4 ). Although tragus measurements are absent from our data set, Kearney (2013a) indicated that the tragus length was slightly less that half the ear length ( 10 mm in holotype and ranging from 10-15 mm in genetically identified type series [n=8]). The tragus of N. stanleyi was previously illustrated under the name N. cf. melckorum ( Van Cakenberghe & Happold 2013a, their Figure 136j) and the distal tip distinctly more rounded than in N. capensis (their Figure 136e). With the exception of ear length, the mean external measurements of N. stanleyi are larger than the N. capensis studied herein, although there is considerable overlap in the range of measurements ( Table 4 View TABLE 4 ).
1 Measurements derived from sequenced specimens.
2 Measurements from specimens obtained in South Africa, Botswana, and Zambia, excluding those from Tanzania. 3 Includes animals from Kersefontein Farm, Western Cape of South Africa described by Roberts (1919) as N. melckorum .
The skull of N. stanleyi is larger and more robust than in specimens of N. capensis and with little overlap in the different measurements. In lateral view, the braincase is low and narrow, similar in general shape to N. capensis , and with inflated occipital and parietal bones creating a weakly developed “helmet” ( Figure 5 View FIGURE 5 ). The zygomatic processes in N. stanleyi are relatively fine, proportionately larger, and more rounded than in N. capensis . In the N. stanleyi holotype (TM 48487), the 2nd upper incisor, which is two-thirds the height of the 1st upper incisor, is unicuspid ( Figure 6 View FIGURE 6 ). In the balance of the specimens allocated to this species based on molecular data, the 2nd upper incisor ranges from one-half to two-thirds the height of the 1st upper incisor and in all cases when the teeth are not too worn (e.g., FMNH 97211), 2nd upper incisor is unicuspid or with a slight bicuspid structure. In general, the 2nd upper incisor in N. capensis shows a slight bicuspid structure. This species has a single upper premolar. The upper and lower molars of N. stanleyi , with respect to the cusp form and general tooth shape, are similar to N. capensis , but in former proportionately larger ( Figure 7 View FIGURE 7 ).
The baculum of N. stanleyi is long, ranging from 2.48–2.55 mm (n = 2), with a distinct ventrally deflected, flattened, roughly triangular short, broad, tip with lateral flanges, a slender shaft, and a base with lobes curving from the lateral edges into the ventral plane ( Figure 2 View FIGURE 2 ).
Description and comparisons. Reports in the literature of external measurements derived from various field collectors display considerable variation associated with different measurement techniques. This problem is further confounded by certain species of Afro-Malagasy bats, in this case small vespertilionids, which include cryptic species showing convergence in morphological characters, further emphasizing the importance in certain cases for identifications to be based on molecular data (e.g., Monadjem et al. 2013; Goodman et al. 2015). This problem is present in the current data set, with Neoromicia capensis showing notable variation in minimum and maximum values for external measurements, presumably outside of collector differences, and whether this is associated with real natural intra-specific variation or composite species complexes is an aspect for future research.
External characters. Kearney (2013a) reviewed species of southern African vespertilionid bats that could be confused with Neoromicia cf. melckorum , named herein as N. stanleyi , based mostly on pelage coloration. Three of these taxa can be easily separated from N. stanleyi using forearm length, which measures on average 37.0 mm (range from 34–39 mm; Table 4 View TABLE 4 ), as compared to an average of 28.1 mm in males and 29.1 mm in females (range from 22–36 mm) of N. somalica , an average of 29.9 mm (range from 27–33 mm) in N. zuluensis , and an average of 31.3 mm (range 29–34 mm) in Hypsugo anchietae ( Kearney 2013a, 2013b; Van Cakenberghe & Happold 2013b). The fourth species, Eptesicus hottentotus has a forearm length averaging 48.2 mm (range 45–54 mm) and, hence, notably larger than N. stanleyi ( Cotterill & Happold 2013) . With an average forearm length of 34.3 mm (range 31– 38 mm), the fifth species, N. capensis , is similar in size to N. stanleyi and this measurement, as well as the other variables presented in Table 4 View TABLE 4 , cannot be used to separate these two species; see cranio-dental characters below. Another species occurring in the region that is similar in size to N. stanleyi is N. rendalli , but it has distinctly lighter pelage coloration and white wings ( Monadjem et al. 2010), which cannot be confused with N. stanleyi .
On the basis of current data, using sequenced individuals, there does not seem to be evidence of notable sexual dimorphism in N. stanleyi . For example, the forearm length of males has an average of 37.0 mm (n = 2) and females of 37.7 mm (n = 5). This same conclusion can be inferred based on measurements of N. cf. melckorum tabulated in Monadjem et al. (2010).
Cranio-dental characters. Dental formula in Neoromicia stanleyi , as in most other members of the genus, is I 2 /3 C 1/1 P 1/2 M 3/3, comprising the adult dentition of 32 teeth and missing the anterior upper premolar. On the basis of cranial measurements, N. stanleyi is distinctly larger than two other members of the capensis group, N. matroka and N. malagasyensis , while the third Malagasy species, N. robertsi is approximately the same size as N.
stanleyi ( Table 5 View TABLE 5 ). However, as these three species are endemic to Madagascar, there is no possible confusion with N. stanleyi based on specimens with locality data. For N. capensis specimens coming from South Africa (including Robert’s [1919] melckorum series), Botswana, and Zambia, the mean measurements of all seven cranial variables used in this study are notably larger in N. stanleyi , and for five measurements these two species show no overlap ( Table 5 View TABLE 5 ). Hence, at least for N. capensis specimens employed in this study, they can be separated from N. stanleyi based on cranial dimensions. However, the situation within what is considered the N. capensis species complex associated with variation in cranio-dental measurements is distinctly more complex (see Discussion).
In general, the skull of N. stanleyi is large and slightly stocky ( Figure 5 View FIGURE 5 ), in many ways proportionately similar to N. robertsi from Madagascar, and distinctly more robust than in all of the specimens of N. capensis examined in this study. In lateral view, the braincase is low and narrow, similar in general shape to N. capensis , and has inflated occipital and parietal bones creating a weakly developed dome shape or has been referred to in the literature on African Neoromicia “helmet”. The sagittal and occipital crests are only slightly developed. In dorsal view, the skulls of N. stanleyi and N. capensis are proportionately similar to one another, with the former being larger than the latter, with the exception of the rostrum being longer and the interorbital region less constricted in N. capensis . The zygomatic processes in N. stanleyi , while being relatively fine structures, are proportionately larger and more rounded than in N. capensis .
GSKL CIL ZYGO POB
N. stanleyi 1 15.2 ± 0.21 14.6 ± 0.26 9.6 ± 0.29 3.7 ± 0.21 t =3.70, df= 11 t =3.69, df= 12 t =3.46, df= 11 t =2.32, df=12 P= 0.001 P= 0.002 P= 0.002 P= 0.02
continued.
MAST PAL MAND
N. stanleyi 1 8.4 ± 0.10 5.5 ± 0.42 10.6 ± 0.32 t =4.00, df= 12 t =2.89, df= 12 t =3.28, df=12 P <0.001 P =0.007 P =0.003
1Measurements derived from sequenced specimens.
2Measurements from specimens obtained in South Africa, Botswana, and Zambia and excludes those from Tanzania. 3 Includes a portion of the type series and other specimens of N. melckorum from Kersefontein Farm, Western Cape of South Africa described by Roberts (1919).
4 Includes only animals from Kersefontein Farm, Western Cape of South Africa, described by Roberts (1919) as N. melckorum .
5 Excludes animals from Kersefontein Farm, Western Cape of South Africa, described by Roberts (1919) as N. melckorum .
As with cranial measurements, the mean values for all five dental variables in N. stanleyi are larger than N. capensis specimens coming from South Africa (including Robert’s [1919] melckorum series), Botswana, and Zambia, but in all cases show broad overlap between these two species in their minimum-maximum range ( Table 6 View TABLE 6 ). In the holotype of N. stanleyi (TM 48487), the 2nd upper incisor, which is two-thirds the height of the 1st upper incisor, is unicuspid ( Figure 6 View FIGURE 6 ). In the balance of the specimens allocated to this species based on molecular analyses, the 2nd upper incisor ranges from one-half to two-thirds the height of the 1st upper incisor and in all cases unicuspid or with a very slight bicuspid structure. In general, the 2nd upper incisor in N. capensis shows an indistinct bicuspid structure, at least in relatively recently erupted adult dentitions ( Kearney 2013a). Another diagnostic character to separate N. stanleyi and N. capensis is the size and robustness of the molar teeth. In the upper molars, the cusp form and general tooth shape are similar, but in N. stanleyi they are proportionately more expanded ( Figure 7 View FIGURE 7 A), although in the lower molars the differences are less pronounced ( Figure 7 View FIGURE 7 B).
t=3.26, df= 12 t =3.89, df= 12 t =3.42, df= 12 t =4.27, df= 10 t =3.10, df=12 P =0.003 P =0.001 P =0.003 P <0.001 P =0.005
1Measurements derived from sequenced specimens.
2Measurements from specimens obtained in South Africa, Botswana, and Zambia and excludes those from Tanzania. 3Includes a portion of the type series and other specimens of N. melckorum from Kersefontein Farm, Western Cape of South Africa described by Roberts (1919).
4Includes only animals from Kersefontein Farm, Western Cape of South Africa, described by Roberts (1919) as N. melckorum .
5Excludes animals from Kersefontein Farm, Western Cape of South Africa, described by Roberts (1919) as N. melckorum .
Male baculum characters. Neoromicia stanleyi has a long baculum ( 2.41–2.55 mm, n = 2), with a distinct, ventrally deflected, flattened, roughly triangular distal tip ( Table 3 View TABLE 3 , Figure 2 View FIGURE 2 ). The tip is short, and broad at the distal edge, which is also slightly concave. There appears to be intraspecific variation in the development of lateral flanges forming distal notches, or possibly these delicate projections were inadvertently cleared during preparation of the baculum, being more apparent in NMZL ES-114/2010 than TM 48487. The slender shaft is transversely concave and evenly wide from proximal to distal ends. In lateral profile the shaft is either straight (TM 48487) or flexed from the vertical plane (NMZL ES-114/2010). The proximal base expands away from the narrower shaft in the dorsal and lateral planes forming flanges that curve around from the lateral edges into the ventral plane creating a concave opening. The proximal edge is even on the dorsal and lateral sides, and slightly ventrally deflected.
Distribution and natural history. Given the confusion that previously existed for bat researchers when handling large vespertilionids in southern Africa between Neoromicia cf. melckorum , named herein as N. stanleyi , and other regional members of this genus, most details on the habits and natural history of N. stanleyi are based on information associated with museum records. In Malawi, it is known to occur in different types of wooded habitats and riverine forests, including Miombo woodlands below 1000 m, as well as lower elevation and drier habitats ( Kearney 2013a).
On the basis of sequenced tissue samples, this species is confirmed to occur in Botswana in the Okavango Delta along the Gomoti River (TM 48484, 48487 [ holotype], FMNH 97211), Zimbabwe in the Mana Pools National Park (TM 41860, 41861, 41862), and Zambia in the Kafue National Park (NMZL ECJS-114/2010) ( Figure 3 View FIGURE 3 ). The Botswana and Zambian specimens were caught in riparian zones. Several other specimens reported in the literature conform to the distinctive characters of this species, but their definitive identifications have not been verified with molecular techniques. On the basis of these records, several other sites can be added to those N. stanleyi is known from, as well as Mozambique to the list of countries ( Monadjem et al. 2010; Kearney 2013a). Some habitat details associated with these specimens include BMNH 66.5460, which was “shot on grassy plain but only 200 yds from Terminalia woodland”. In a study of the impact of habitat disturbance in close vicinity to the Mana Pools National Park, the species identified as N. melckorum , which is presumed to be N. stanleyi , was only found, albeit rarely, in intact woodlands ( Fenton et al. 1998). Most of the localities in the northern part of Kruger National Park in South Africa, from which N. stanleyi has been tentatively identified, were also within riparian vegetation along the Levuvhu River (see locality information in Fenton and Rautenbach (1986) and Aldridge and Rautenbach [1987]).
Few details are available on the diet of N. stanleyi , but based on a limited sample it is known to feed on (in order of importance) Coleoptera, Hymenoptera, Dictyoptera (particularly mantids), and Lepidoptera ( Fenton et al. 1998). A number of aspects on life history traits of N. stanleyi are difficult to separate from details reported in the literature for “ N. melckorum ”, which is now considered a synonym of N. capensis , or other Neoromicia spp. Some details on the echolocation calls for what might be N. stanleyi are presented by Kearney (2013a).
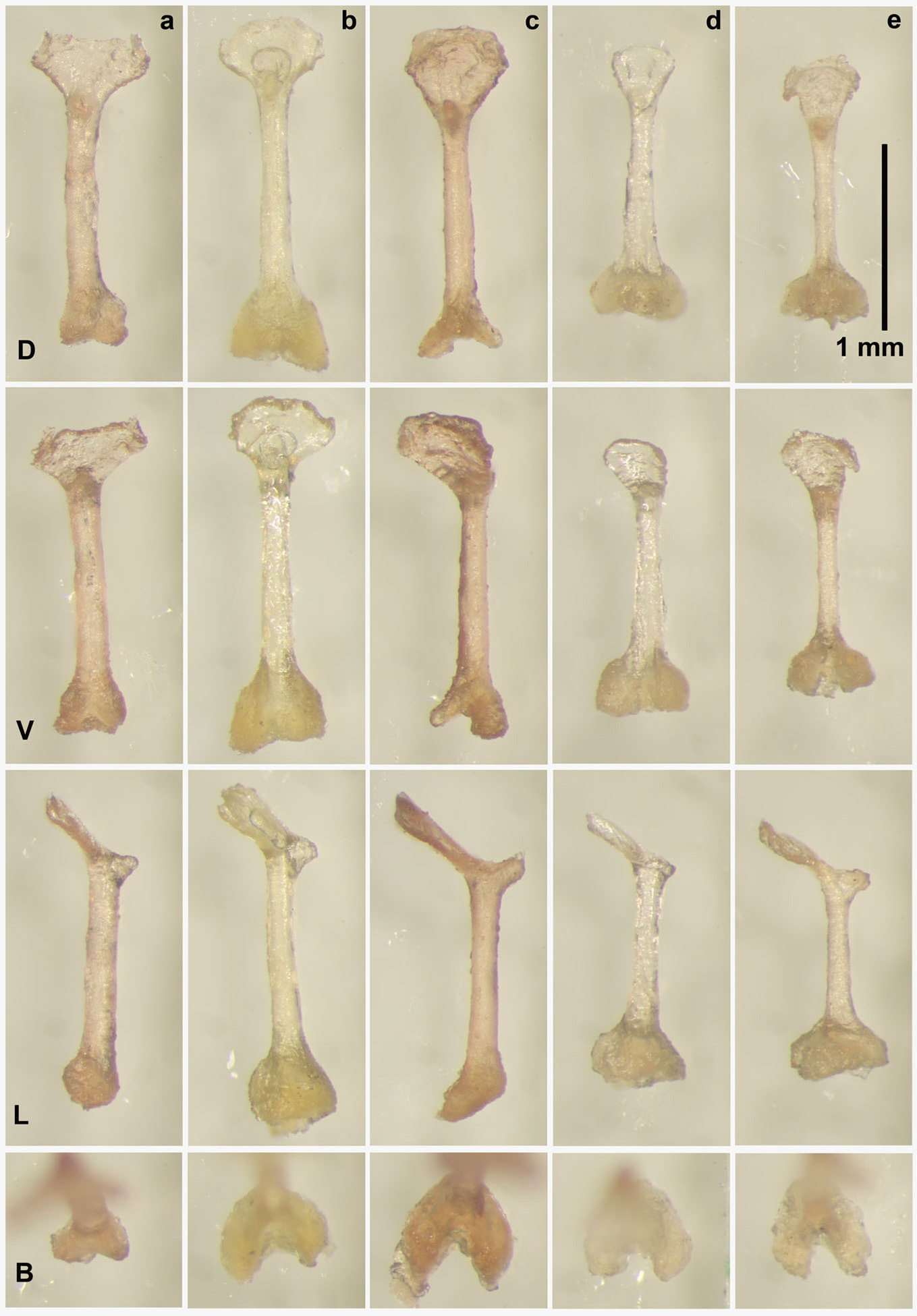
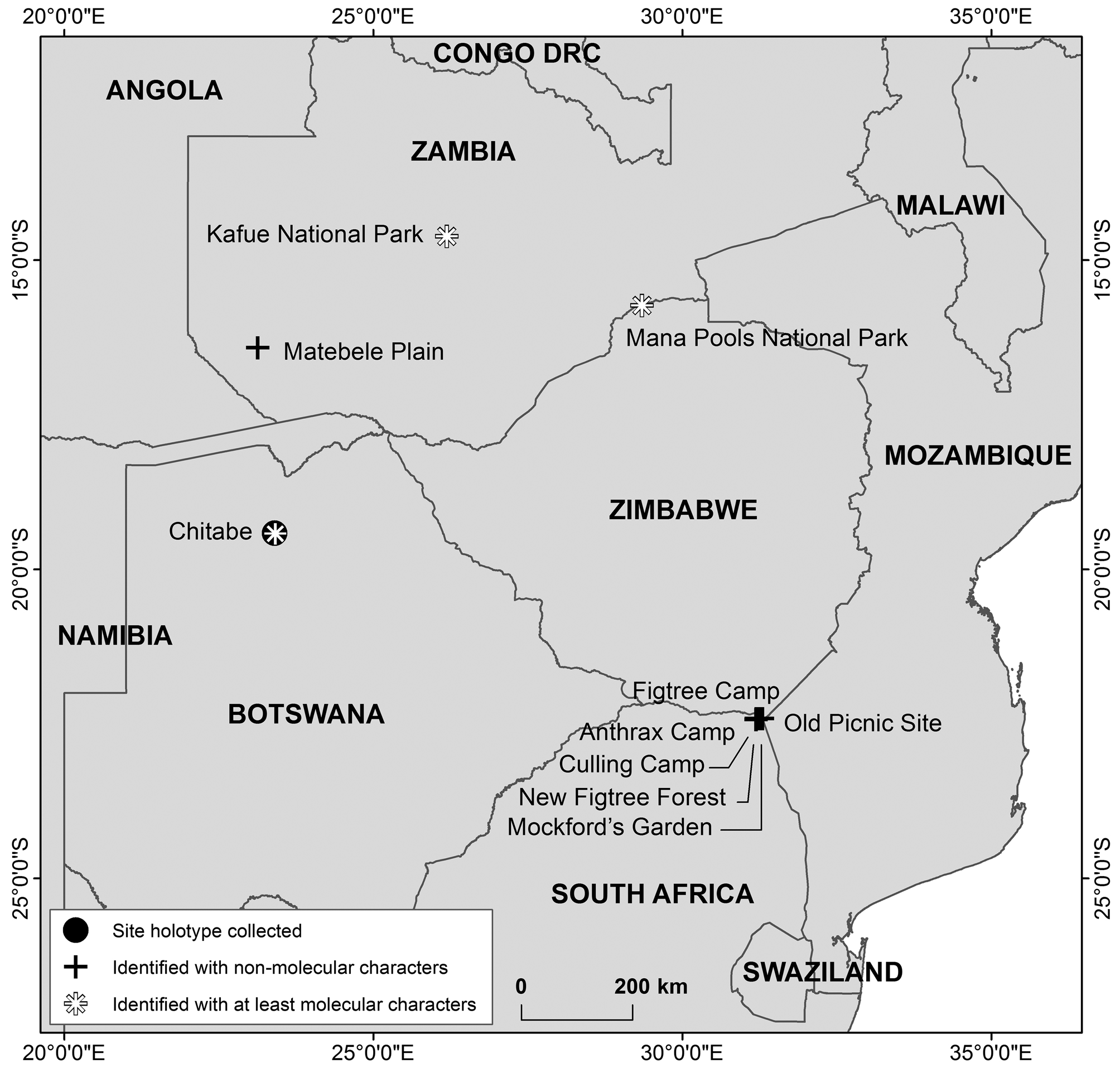

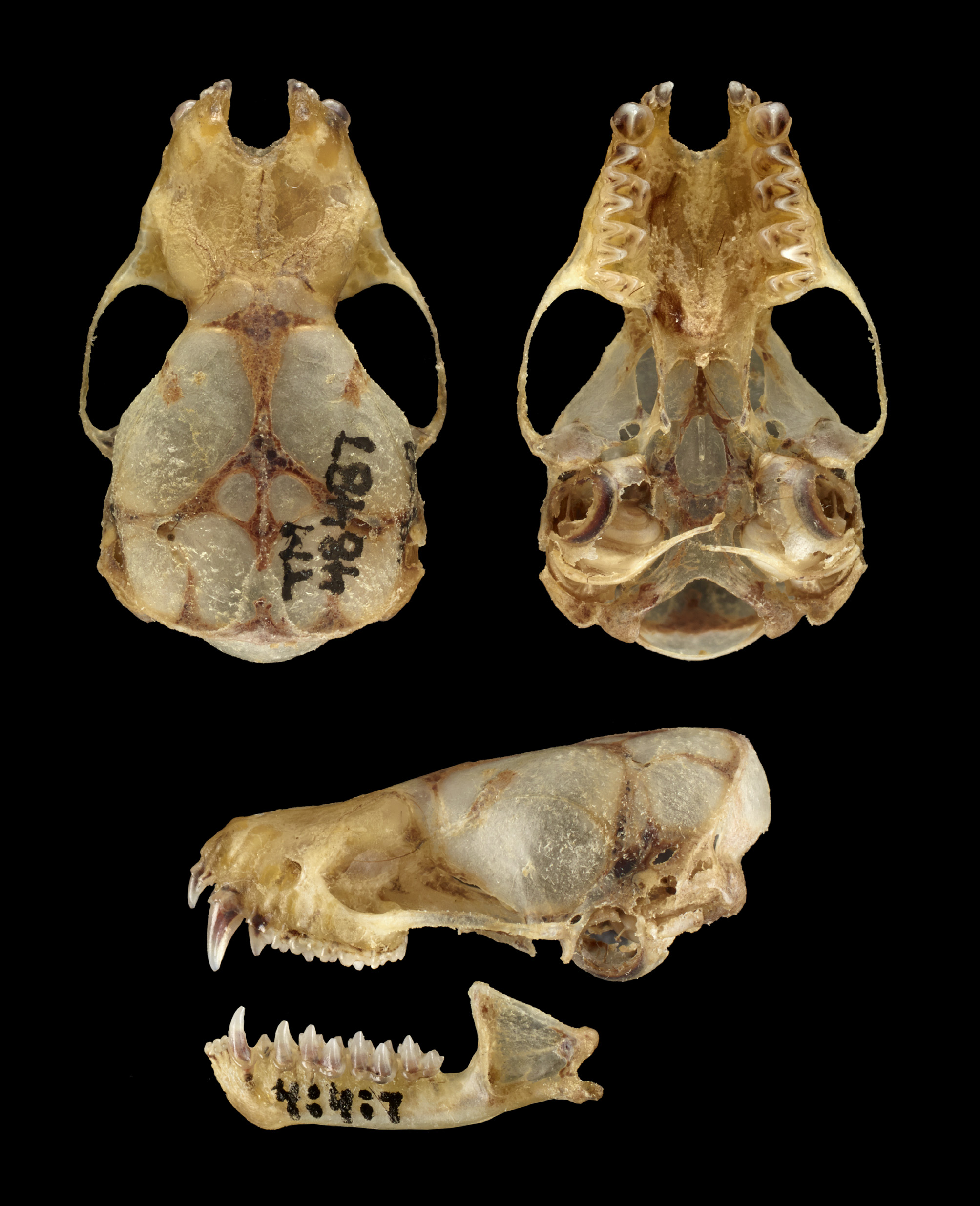
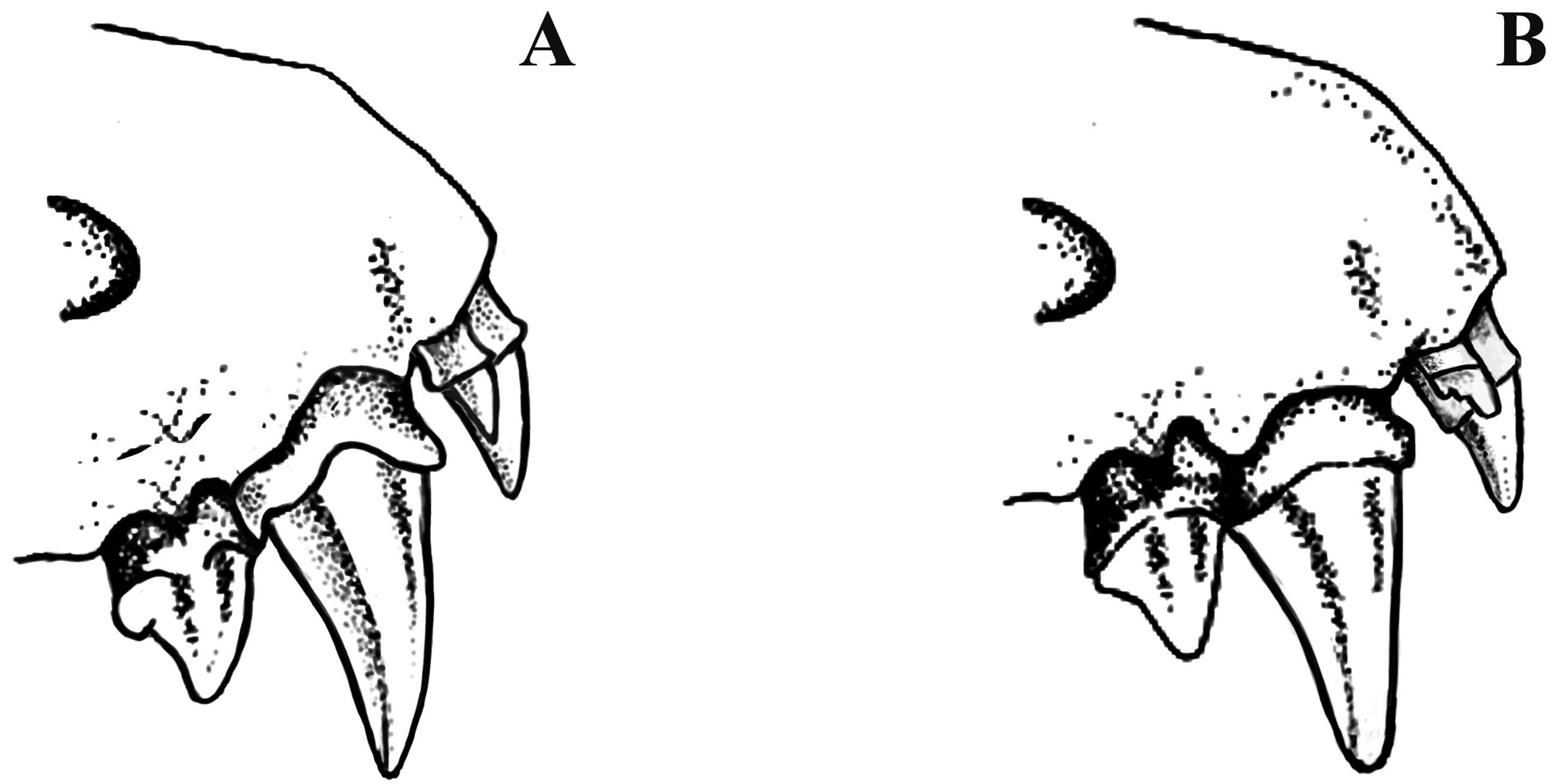
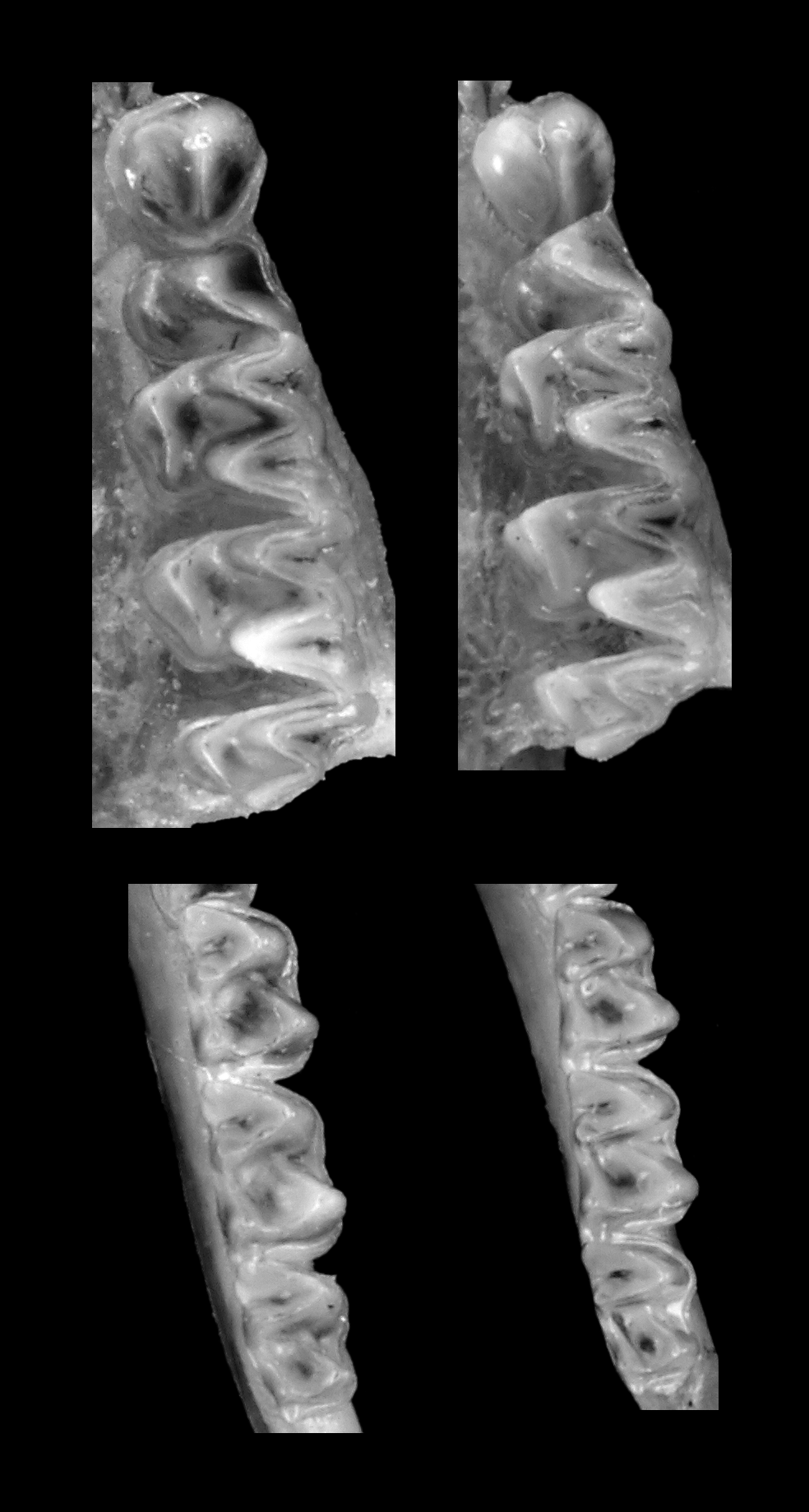
TABLE 4. External measurements (in millimeters) and mass (in grams) of Neoromicia stanleyi sp. nov., N. robertsi, N. matroka, N. malagasyensis, and N. capensis. Measurements presented as mean ± standard deviation, minimum and maximum measurements, and number of specimens. Given small sample sizes, aspects of sexual dimorphism for the taxa presented here have not been taken into account. In the case of N. stanleyi and N. capensis, the notably high standard deviation values and the broad range of minimum and maximum measurement values are presumably an artifact of different measurement techniques used by various field collectors.
| Total length | Tail length | Hindfoot length | Tragus length | |
|---|---|---|---|---|
| N. stanleyi 1 | 93.3 ± 6.42 83–101, n=8 | 39.7 ± 4.46 34–48, n=8 | 7.3 ± 0.76 6.0–8.5, n=8 | - |
| TM 48487, holotype | 92 | 48 | 7 | - |
| N. robertsi | 91.0 ± 4.06 84 – 94, n=5 | 34.6 ± 2.07 31 – 36, n=5 | 5.4 ± 0.55 5 – 6, n=5 | 7.2 ± 1.10 6 – 8, n=5 |
| N. matroka 1 | 82.1 ± 3.18 77 – 86, n=12 | 32.2 ± 2.76 27 – 36, n=12 | 4.6 ± 0.51 4 – 5, n=12 | 6.8 ± 0.50 6 – 8, n=12 |
| N. malagasyensis 1 | 81.3 ± 1.15 80 – 82, n=3 | 36.0 ± 1.00 35 – 37, n=3 | 4.8 ± 0.50 4 – 5, n=4 | 6.5 ± 0.58 6 – 7, n=4 |
| N. capensis 1,2,3 | 87.7 ± 4.36 81 – 96, n=12 | 37.1 ± 4.85 32 – 45, n=15 | 7.1 ± 1.07 6 – 9, n=15 | 6.7 ± 0.58 6 – 7, n=3 |
| continued. |
TABLE 5. Cranial measurements (in millimeters) of adult Neoromicia stanleyi sp. nov., N. robertsi, N. matroka, N. malagasyensis, N. capensis, and N. melckorum. Measurements presented as mean ± standard deviation, minimum and maximum measurements, and number of specimens. See Methods for an explanation of variable acronyms. Given small sample sizes for most taxa, aspects of sexual dimorphism have not been taken into account. The T-test statistics at the foot of the table are to compare measurement differences between Roberts’ N. melckorum and specimens of N. capensis obtained in other areas of South Africa, Botswana, and Zambia.
| 14.8–15.4, n=7 | 14.3–15.0, n=7 | 9.3–10.0, n=6 | 3.5–4.1, n=7 | |
|---|---|---|---|---|
| TM 48487, holotype | 15.1 | 14.3 | 9.3 | 3.7 |
| N. robertsi | 14.7 ± 0.19 14.4 – 14.8, n=4 | 14.1 ± 0.05 14.1 – 14.2, n=4 | 9.9 ± 0.37 9.5 – 10.4, n=4 | 4.0 ± 0.24 3.6 – 4.1, n=4 |
| N. matroka 1 | 13.1 ± 0.33 12.4 – 13.4, n=12 | 12.6 ± 0.35 12.0 – 13.0, n=12 | 8.8 ± 0.28 8.4 – 9.2, n=9 | 3.5 ± 0.14 3.3 – 3.8, n=12 |
| N. malagasyensis 1 | 12.6 ± 0.38 12.4 – 13.3, n=5 | 12.0 ± 0.40 11.7 – 12.7, n=5 | 8.5 ± 0.26 8.3 – 8.8, n=5 | 3.4 ± 0.18 3.2 – 3.7, n=5 |
| N. capensis 1,2,3 | 14.0 ± 0.33 13.4 – 14.3, n=8 | 13.6 ± 0.37 13.1 – 14.1, n=8 | 8.8 ± 0.28 8.5 – 9.2, n=8 | 3.5 ± 0.05 3.4 – 3.5, n=8 |
| “ N. melckorum ” 4 | 15.2 ± 0.46 14.7 – 15.5, n=3 | 14.7 ± 0.52 13.9 – 15.0, n=4 | 9.5 ± 0.17 9.4 – 9.7, n=3 | 3.7 ± 0.13 3.5 – 3.8, n=4 |
| N. capensis 5 | 14.1 ± 0.46 13.4 – 15.1, n=10 | 13.6 ± 0.47 13.1 – 14.5, n=10 | 8.8 ± 0.32 8.5 – 9.4, n=10 | 3.5 ± 0.15 3.1 – 3.7, n=10 |
TABLE 6. Dental measurements (in millimeters) of adult Neoromicia stanleyi sp. nov., N. robertsi, N. matroka, N. malagasyensis, and N. capensis. Measurements presented as mean ± standard deviation, minimum and maximum measurements, and number of specimens. See Methods for an explanation of variable acronyms. Given small sample sizes for most taxa, aspects of sexual dimorphism have not been taken into account. The T-test statistics at the foot of the table are to compare measurement differences between Roberts’ N. melckorum and specimens of N. capensis obtained in other areas of South Africa, Botswana, and Zambia.
| I-M3 | C-M3 | C-C | M3-M3 | i-m3 | |
|---|---|---|---|---|---|
| N. stanleyi 1 | 5.9 ± 0.15 5.7–6.1, n=7 | 5.1 ± 0.13 4.9–5.3, n=7 | 4.6 ± 0.20 4.3–4.8, n=7 | 6.1 ± 0.17 5.8–6.3, n=7 | 5.7 ± 0.33 5.4–5.9, n=6 |
| TM 48487, holotype | 5.8 | 5.0 | 4.4 | 5.8 | 5.5 |
| N. robertsi | 6.1 ± 0.10 6.0 – 6.2, n=4 | 5.4 ± 0.13 5.2 – 5.5, n=4 | 4.9 ± 0.17 4.6 – 5.0, n=4 | 6.5 ± 0.10 6.4 – 6.6, n=4 | 5.7 ± 0.10 5.5 – 5.7, n=4 |
| N. matroka 1 | 5.2 ± 0.17 5.0 – 5.4, n=12 | 4.5 ± 0.17 4.2 – 4.8, n=12 | 4.1 ± 0.17 3.9 – 4.4, n=12 | 5.6 ± 0.20 5.3 – 5.9, n=12 | 4.8 ± 0.18 4.5 – 5.2, n=12 |
| N. malagasyensis 1 | 4.8 ± 0.18 4.5 – 5.0, n=5 | 4.2 ± 0.13 4.1 – 4.4, n=5 | 3.8 ± 0.16 3.6 – 4.0, n=5 | 5.2 ± 0.30 5.0 – 5.7, n=5 | 4.6 ± 0.11 4.5 – 4.7, n=5 |
| N. capensis 1,2,3 | 5.6 ± 0.14 5.4 – 5.7, n=8 | 4.8 ± 0.13 4.7 – 5.0, n=8 | 4.4 ± 0.12 4.2 – 4.5, n=8 | 5.7 ± 0.17 5.6 – 6.0, n=8 | 5.3 ± 0.16 5.1 – 5.5, n=8 |
| “ N. melckorum ” 4 | 6.1 ± 0.19 5.8 – 6.2, n=4 | 5.3 ± 0.13 5.1 – 5.4, n=4 | 4.7 ± 0.19 4.4 – 4.8, n=4 | 6.2 ± 0.22 5.9 – 6.4, n=4 | 5.8 ± 0.19 5.5 – 5.9, n=4 |
| N. capensis 5 | 5.6 ± 0.26 5.3 – 6.2, n=10 | 4.8 ± 0.21 4.6 – 5.3, n=10 | 4.3 ± 0.16 4.0 – 4.5, n=10 | 5.7 ± 0.17 5.6 – 6.0, n=8 | 5.3 ± 0.28 4.9 – 5.9, n=10 |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Neoromicia stanleyi
| Goodman, Steven M., Kearney, Teresa, Ratsimbazafy, Malalatiana Michèle & Hassanin, Alexandre 2017 |
Pipistrellus cf. melckorum
| Kearney 2013 |
Neoromicia cf. melckorum Monadjem et al ., 2010
| Monadjem et al. 2010 |
Neoromicia melckorum
| Simmons 2005 |
Eptesicus ( Neoromicia ) melckorum
| Koopman 1993 |
Pipistrellus cf. melckorum
| Rautenbach et al. 1993 |
Pipistrellus ( Neoromicia ) melckorum
| Hill & Harrison 1987 |
Eptesicus melckorum
| Hayman & Hill 1971 |