
Glaphyropoma spinosum, Bichuette & Pinna & Trajano, 2008
|
publication ID |
https://doi.org/ 10.1590/S1679-62252008000300002 |
|
persistent identifier |
https://treatment.plazi.org/id/03A087EB-582B-9F20-34D4-FEEBFCA9BE16 |
|
treatment provided by |
Carolina |
|
scientific name |
Glaphyropoma spinosum |
| status |
sp. nov. |
Glaphyropoma spinosum View in CoL , new species
Figs. 1-6 View Fig View Fig View Fig View Fig View Fig View Fig
Holotype. MZUSP 99742 View Materials , 58.2 mm SL, Brazil, State of Bahia, Chapada Diamantina, Município de Andaraí, Povoado de Igatu, Gruna dos Torras , rio Paraguaçu drainage, 12º52’40.4”S 41º18’43.3”W, 20 Jan 2006, M. E. Bichuette, F. D. Passos & R. Borowsky. GoogleMaps
Paratypes. MZUSP 99743 View Materials , 4 View Materials (1 c&s), 34.9-45.6 mm SL, same locality as the holotype, 2 Jan 2007, M. E. Bichuette, F. D. Passos GoogleMaps & T. L. C. Scatolini .
Diagnosis. Distinguished from all other copionodontine catfishes by the presence of opercular odontodes. Further distinguished from its only congener, G. rodriguesi , by the reduction of dark integumentary pigmentation.
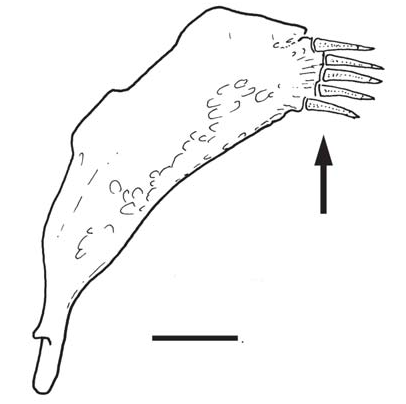
Description. Morphometric data for holotype and paratypes presented in Table 1. For general aspect refer to Fig. 1 View Fig . Body elongate, depressed in head and anterior trunk region, crosssection roughly circular from middle of trunk to origin of dorsal fin, gradually more compressed towards tail. Caudal peduncle region strongly compressed. Dorsal profile of trunk near dorsal fin nearly parallel to longitudinal axis of body, gently convex anteriorly and continuous with dorsal profile of head. Dorsal profile of body posterior to dorsal fin straight to base of caudal fin. Ventral profile of head straight, continuing in gently convex curve to pelvic-fin insertion. Ventral profile of body from origin of pelvic fin to origin of caudal peduncle nearly straight, slightly concave along caudal peduncle. Myotomes not evident through skin, except near caudal fin base.
Integument thick, especially on back and sides of trunk, forming folds on sides of trunk in large specimens, thinnest on abdominal region. Bases and proximal portions of all fins covered with thick layer of integument, most notably on dorsal and anal fins. Skin papillae small, most concentrated on ventral surface of head and barbels.
Head wide and depressed, wider than long in large specimens (holotype and larger paratype; Fig. 2 View Fig ), and slightly narrower in smaller specimens. Shape of head in large specimens nearly semicircular in dorsal view.Anterior part of lateral margin of head expanded over base of maxillary barbel. Eyes very small, without free orbital rim, almost entirely dorsal on head and partly sunk in integument. Lens visible through integument. Eyes located slightly closer to tip of snout than to dorsal margin of branchial membrane, and nearly equidistant between midline and lateral margin of head.
Anterior naris extended anterolaterally by tube of integument, continuous with nasal barbel. Posterior naris closer to eye than to anterior naris, partially closed by two thin flaps of skin not forming continuous tube. Posterior nares closer together than anterior ones.
Mouth inferior, located close to anterior margin of snout, not strongly flattened ventrally ( Fig. 3 View Fig ). Upper and lower lips narrow in ventral view, the latter not divided in lateral halves by ridge. Posterior limit of lower lip not well-defined, mostly continuous with mental region ( Fig. 3 View Fig ). Angle of mouth with fleshy outgrowth just posterior to rictal barbel.
Most teeth in first row of premaxilla and dentary distally expanded and flat, symmetrically spatulate and not overlapping distally. Fourteen or fifteen teeth in first dentary row extending nearly entire length of bone to base of coronoid process. Teeth progressively smaller and less spatulate laterally, with 3 or 4 lateralmost teeth conical. Eleven or twelve teeth on outer row of premaxilla, similar in morphology to those on dentary and with 2 or 3 lateralmost ones conical. Distal portions of all teeth darkly-pigmented, more markedly so in spatulate ones. Second row of premaxillary and dentary teeth much less markedly spatulate than those in first row. Teeth of subsequent rows mostly conical, with a few slightly compressed at tip.
Branchial membranes thick, narrowly attached to isthmus anteriorly. Branchiostegal rays seven, not visible through skin in alcohol-preserved specimens.
Barbels long and slender, with visible internal cores, their length variable bilaterally and among specimens. Maxillary barbel reaching between middle and posterior margin of pectoral-fin base. Rictal barbel extending to middle of interopercular patch of odontodes. Nasal barbel reaching anterior margin of pectoral-fin base or its middle.
Interopercle with two rows of odontodes ( Fig. 3 View Fig ), ventral (large) and dorsoposterior (small). Ventral patch of odontodes extending along posterior part of ventral margin of head, its anterior point separated from lower lip by distance roughly equivalent to mouth width. Posterior point of patch nearly reaching vertical through anterior margin of pectoral-fin base. Odontodes light brown on distal half, disposed in two irregular rows, those on inner (posterior) row larger. Odontodes increasing in size posteriorly, with largest ones located at posterior tip of inner row. Largest odontodes slightly flattened and expanded distally, round at tip and bent at midlength. Eight or nine odontodes in inner row of ventral patch. Dorsal patch much smaller than ventral one, located laterally on posterior portion of head and with four to six small conical odontodes disposed in single curved row. Odontodes of dorsal patch almost entirelly hidden in integument, visible externally by their individual skin clefts and darkly-pigmented tips. Opercle with small odontode patch located slightly anterior to vertical through origin of pectoral fin. Five or six small conical opercu- lar odontodes arranged in single row, with their axes oriented posteroventrally ( Fig. 4 View Fig ). Opercular patch of odontodes underlain by small fold of integument and mostly hidden in skin, with only tips of individual odontodes exposed.
Lateral-line canal externally visible, extending midlaterally in four to seven interrupted segments from head to, at most, vertical through origin of pelvic fins. Lateral-line ossicles absent. Infraorbital branch of latero-sensory canal system complete, detectable externally by its three pores or in outline in emaciated specimens. Infraorbital canal leaving neurocranium exclusively through sphenotic. Infraorbital bones absent except for first one (lacrimal).
Pectoral fin elongated, roundish-triangular in shape when expanded, its insertion slightly anterior to dorsal limit of branchial membrane ( Figs. 1 View Fig , 2 View Fig ). Muscular base of pectoral fin broad. Pectoral-fin rays i+10, with first (unbranched) ray strongly curved posteriorly (more so along proximal half) and approximately same length as remaining ones, not prolonged as filament.
Pelvic fin medium size with round distal margin. Origin of pelvic fin closer to tip of snout than to base of caudal fin, at vertical through origin of dorsal fin. Pelvic-fin rays i+5 (one paratype) or i+6 (remaining specimens), with first ray (unbranched) curved and shorter than all succeeding rays but for last one. Pelvic splint absent.
Dorsal-fin origin equidistant between base of caudal fin and tip of snout, with ii+7 (most specimens) or ii+8 (c&s paratype) rays. Anal fin larger than dorsal, with broadly convex profile, its origin slightly closer to medial point of branchial membrane than to posterior margin of caudal fin.Anal-fin rays iii+9, ii+9 or ii+10 (holotype), plus two anterior procurrent rays.
Caudal fin shape variable, ranging from truncate, to truncate with round edges, emarginate or slightly concave. Posterior profile oblique, resulting in upper lobe longer than lower one. Principal caudal-fin rays i+7 and 8+i, procurrent rays weakly developed, five dorsally and six or seven ventrally.
Adipose fin large and conspicuous, extending for nearly entire region between dorsal and caudal fins.Anterior origin of adipose fin gently rising from immediately posterior to dorsal fin, reaching greatest depth on its posterior half and posteriorly continuous with caudal fin. Limit between two fins not well-defined, located in region with concave profile. Entire adipose fin covered with nearly continuous layer of large adipose tissue.
Vertebrae 30 or 31, first dorsal-fin pterygiophore immediately posterior to neural spine of vertebra 7; first anal-fin pterygiophore immediately posterior to hemal spine of vertebra 13. Nine pleural ribs.
Pigmentation in alcohol. Dark coloration faint, most specimens white in superficial aspect, although none lacking dark integumentary pigment altogether. Dark chromatophores mostly concentrated on dorsal part of head and dorsum, uniformly scattered, not forming definite pattern. Densest concentration along middle of dorsum, forming broad dark middorsal band in some specimens, abruptly fainter anteriorly at edge of neurocranium and gradually fainter posteriorly, disappearing entirely posterior to dorsal fin ( Fig. 2 View Fig ). Dorsal sur- face of head with uniform scattering of dark chromatophores, densest on upper half of sides. Some concentration of dark pigment also immediately anterior to anterior nasal barbel. Cranial fontanels visible in white, due to concentration of adipose tissue. Barbels lacking dark pigment except for faint fields of dark chromatophores at base of nasal and, sometimes, maxillary barbel. Opercular and interopercular odontodes darkly pigmented at their distal portions, with their protruding portions clearly visible against pale background integument ( Figs. 1 View Fig , 3 View Fig ). Fins white. Ventral part of body and head lacking dark integumentary pigment ( Fig. 3 View Fig ).
Pigmentation in life. Pinkish on dorsal portion of body and lateral portion of head, showing an iridescent aspect. Cranial fontanel white. White coloration on lateral and ventral portions of body, due to concentration of adipose tissue. Eyes black ( Fig. 5 View Fig ).
Etymology. The name spinosum is Latin for prickly, spiny, in allusion to the presence of opercular odontodes in this species, unique within the Copionodontinae .
Ecological data. The type locality is a quartzitic cave situated within the limits of the Chapada Diamantina National Park, in the Serra do Sincorá, one of the main components of the Chapada Diamantina, inside the limits of the Chapada Diamantina National Park. The local terrain is composed of arenites, siltites, argilites and conglomerates. Caves in this area are formed by the erosion of soft rock components by rainwater penetrating through surface cracks and leaving spaces delimited by the harder components. The water depth in the creek where the catfish live varies mostly between 10 and 50 cm, with moderate current and some spotty accummulation of plant debris where fish are found foraging. Physico-chemico water variables in January 2007 were pH 4.1; temperature 23.0ºC; conductivity 0.012 PS/cm; dissolved oxygen 6.3 mg /l.
Preliminary population data based on visual censuses were obtained in January and July, 2007, corresponding respectively to the rainy and dry seasons in the area. In a 150m of long stream reach and 1m of width (in average), 61 individuals were visually counted in January and 59 in July of 2007, and respectively population densities of 0.41 ind.m-2 (Jan 2007) and 0.39 ind.m-2 (Jul 2007), indicating little if any seasonal oscillations in population size. Most individuals were concentrated in quiet isolated pools ( Fig. 6 View Fig ). The subterranean course of the creek extends for at least another 2,000 meters downstream, where, apparently the population is distributed (A. Auler & L. Zogbi, pers. com.). Most of the caves in the upper sector of Chapada Diamantina were heavily impacted by diamond mining in the past, an activity which extended to the early 1990’s and, in a residual clandestine fashion, continues to the present. This impact is visible in the blocks of rocks and remains of the miners inside the caves (lamps, bottles).
Additional copionodontine specimens found inside the cave may represent two other species that co-occur with G. spinosum . One of them is clearly related to Copionodon orthiocarinatus , and the other possible to C. pecten . In both cases the individuals found in the cave are troglomorphic, with a visible reduction of dark pigmentation, in some specimens, also of eyes in some specimens. A few individuals seem to be intermediates of Glaphyropoma and Copionodon catfish. Proper resolution of the taxonomic significance of these forms will require more detailed studies. The sympatric ocurrence of three copionodontine species has been recorded in the type locality of G. rodriguesi , C. pecten and C. orthiocarinatus , therefore it is not unexpected for this to also occur in the cave.




| R |
Departamento de Geologia, Universidad de Chile |
| T |
Tavera, Department of Geology and Geophysics |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |