
Sheldonia fingolandensis, Herbert, 2017
|
publication ID |
https://doi.org/ 10.5852/ejt.2017.309 |
|
publication LSID |
lsid:zoobank.org:pub:1E8FE779-D6E7-428E-9538-5E5F8ECFB271 |
|
DOI |
https://doi.org/10.5281/zenodo.3846864 |
|
persistent identifier |
https://treatment.plazi.org/id/039F87A5-676D-FFB3-4D29-FD94FE6AFCD7 |
|
treatment provided by |
Carolina |
|
scientific name |
Sheldonia fingolandensis |
| status |
sp. nov. |
Sheldonia fingolandensis View in CoL sp. nov.
urn:lsid:zoobank.org:act:8620960E-0BBC-49D2-9CF6-69D028EDB052
Figs 15 View Fig , 31–34 View Fig View Fig View Fig View Fig
Diagnosis
Characterised by the axially striped colour pattern, large tumescent shell, finely punctate protoconch sculpture and arboreal habit.
Etymology
From Fingoland, the territory between the Kei and Mbhashe rivers, E Cape, inhabited by the Fingo people (amaFengu) after their flight south to escape the brutality of King Shaka in the early 1800s.
Material examined
Holotype
SOUTH AFRICA: E Cape, Mbhashe River valley, Collywobbles , overlooking vulture colony, 32.00396° S, 28.59530° E, 585 m, sparse valley thicket invaded with Lantana , living on aloes, D. Herbert and L. Davis leg., 20 Feb. 2006 ( NMSA W9688/T3383 , dry shell).
GoogleMapsParatypes (listed north to south, all E Cape)
SOUTH AFRICA: Mbhashe River valley, near N2 bridge, 31.92461° S, 28.45113° E, 470 m, riverine thicket, living on aloe leaves, A. Moussalli and D. Stuart-Fox leg., 23 Nov. 2005 ( NMSA W4169/T3385, six dry shells with six bodies and eight whole specimens in ethanol); Mbhashe River valley, near N2 bridge, 31.92461° S, 28.45113° E, 470 m, riverine thicket, dead in leaf-litter and under fallen aloes, A. Moussalli and D. Stuart-Fox leg., 23 Nov. 2005 ( NMSA W5373/T3386, two dry shells); same data as holotype ( NHMUK 20160245, one dry shell; NMSA W4075/T3384, seven dry shells with bodies in ethanol; RMNH.5004187, one dry shell); Mbhashe River valley, Collywobbles, slope below carpark on neck, north-facing, valley thicket, under leaves of Aloe ferox , 23.00436° S, 28.58169° E, 510 m, M. Bursey leg., 20 Feb. 2006 ( ELM W2927/T164, five dry shells with bodies in ethanol); Mbhashe River valley, Collywobbles, beside thatched picnic site, 32.004417° S, 28.58172° E, 510 m, valley thicket, living under green leaves of aloes, D. Herbert and L. Davis leg., 20 Feb. 2006 ( NMSA W3976/ T3387, 20 dry shells with 14 bodies in ethanol); Kei River valley, NNE of Stutterheim, Wedgeley Farm, Shed Camp, 32.28986° S, 27.54674° E, 835 m, valley thicket, D. Herbert and L. Davis leg., 4 Mar. 2007 ( NMSA W5345/T3388, one dry shell); Sihota, west bank of Kei River, approx. 2.5 km upstream of Kei Bridge, below Moordenaars Kop, 32.50445° S, 27.94945° E, 200 m, under Aloe ferox leaves, M. Cole leg., 9 Jan. 2008 ( ELM W3263/T165, one dry shell with body in ethanol; NMSA W6386/T3389, six dry shells with two bodies in ethanol).
Other material (listed north to south, all E Cape)
SOUTH AFRICA: Kei Pass, 32.49364° S, 27.99175° E, 282 m, valley thicket, dead under rocks, A. Moussalli and D. Stuart-Fox leg., 23 Nov. 2005 ( NMSA W5486); Mncotsho, Buffalo River valley,
32.917° S, 27.583° E, 360 m, under scattered bushes (esp. Coddia rudis) in grassland near beacon, M. Cole and C. Vernon leg., 6 Nov. 2007 ( NMSA P0630).
Description
SHELL ( Fig. 31 View Fig ). Subglobose to globose, spire prominent; periphery at or close to mid-whorl, evenly rounded; H:D 0.72–0.83 (N=9); suture rather distinctly indented, inserting just above periphery; thin, translucent straw-brown when fresh, sometimes with a greenish tint and with conspicuous pattern of unevenly spaced, opaque, pale buff-white axial lines and stripes; apical surface satin-like, base more glossy. Protoconch diameter 2.3–2.7 mm (N=6); junction with teleoconch sometimes clear, sometimes weakly marked; sculpture very fine, comprising numerous close-set, microscopic spiral lirae, these initially composed of microscopic punctae ( Fig. 31E View Fig ); punctae also with an axial alignment, producing a secondary collabral pattern; latter half whorl of protoconch with spiral lirae only (no longer punctate). Teleoconch of up to almost 3.0 whorls; early part of first whorl with fine spiral sculpture similar to that on protoconch, but finer; remainder of shell with only finest traces of close-set, microscopic lirae, producing satin-like shell surface; an axial component not evident in microsculpture, but uneven growth-lines frequent and appearing to coincide with axial variation in colour pattern; occasional sparse malleation also evident, but insignificant; base more glossy, microsculpture becoming obsolete below periphery. Umbilicus completely absent in adult specimens, edge of columella lip reflected and slightly thickened with whitish callus, running into axis of coiling rather than fusing with parietal region; juvenile and subadult specimens often minutely rimate, columella lip lacking callus thickening and umbilicus remaining as a narrow open channel inside reflected lip. Aperture subcircular, but indented by penultimate whorl in parietal region. Diameter up to 20.9 mm; holotype, diameter 18.7 mm, height 15.0 mm.
LIVING ANIMAL ( Fig. 32 View Fig ). Head-foot mostly pale yellowish-buff, neck and mantle edge somewhat darker orange-buff, tentacles similarly pale, but caudal appendage slightly darker; coloration of spire viscera generally not visible through shell, but cream blotches sometimes evident overlying darker digestive gland.
RADULA ( Fig. 33 View Fig ). Formula R+17+1+(60–70); rachidian tricuspid; laterals essentially bicuspid with a mesocone and strong basal ectocone, but also with a minute endocone on side of mesocone (scarcely more than a low bump); laterals followed by one intermediary tooth and then a long series of marginals; marginals curved, bearing a large terminal cusp with a smaller subterminal cusp on outer (concave) margin, additional irregular cusps may be present on shaft, but these normally obscured by overlapping outer neighbouring tooth; marginals progressively decreasing in size toward edge of radula, but otherwise morphologically similar.
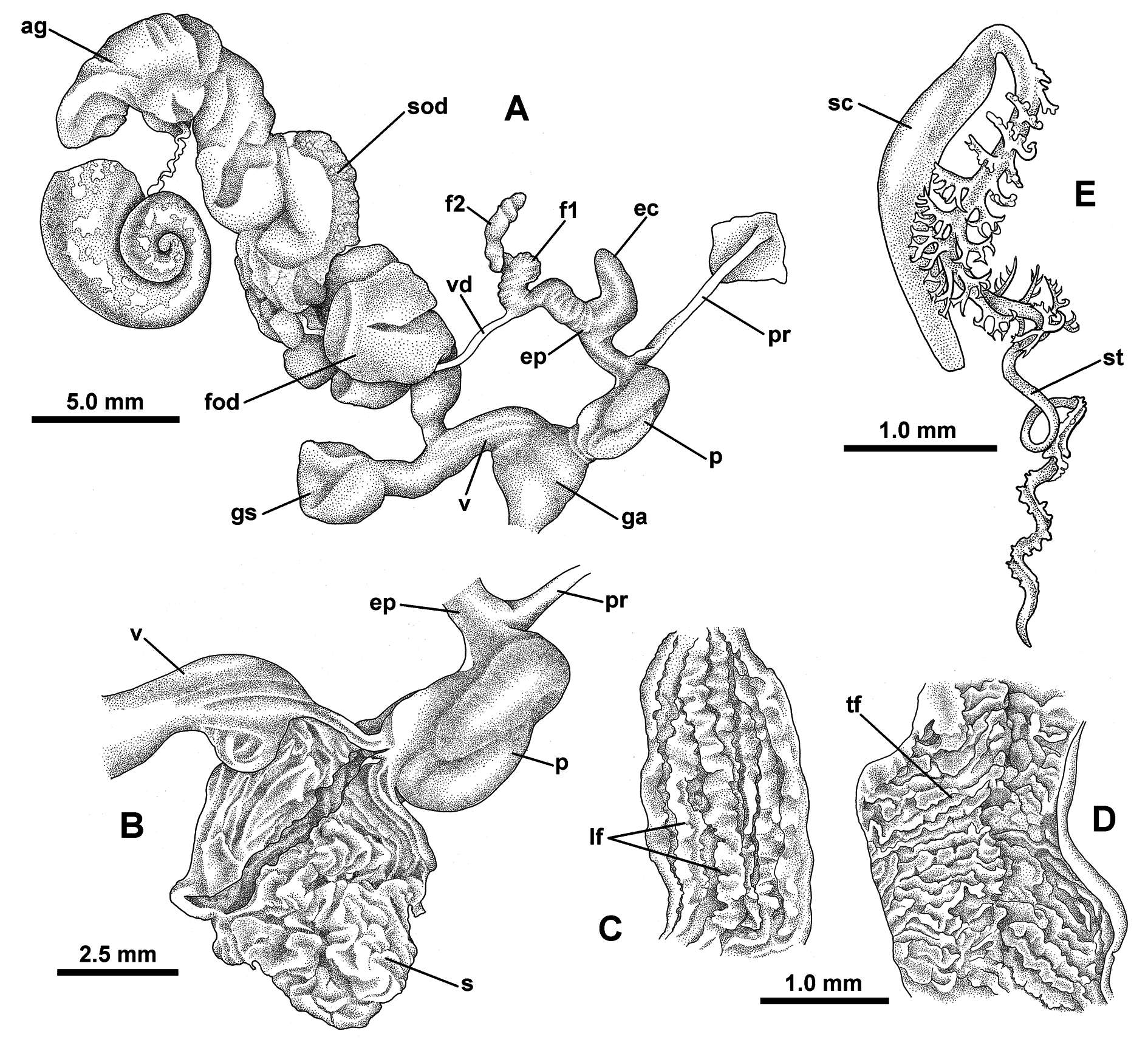
DISTAL GENITALIA ( Fig. 34 View Fig A–D). Penis elongate and cylindrical, but conspicuously looped upon itself and surrounded by a thin sheath, 2–3 times longer when removed from sheath; retractor muscle attached to
penis apex; basal half of penis relatively thin-walled, its lumen lined with close-set, irregularly sinuous transverse folds ( Fig. 34D View Fig ); apical half of penis with thicker wall, lumen lined by longitudinal ridges, these largely smooth near junction with epiphallus, but progressively stronger and more zig-zag toward mid-region of penis ( Fig. 34C View Fig ). Epiphallus short, with a stout, recurved caecum; caecum arising close to middle of epiphallus; lumen of caecum coarsely and irregularly papillate. Flagellum well developed, basal portion (f1) broader and with distinct transverse internal structure; distal portion (f2) longer and more slender, twisted into a spiral of several revolutions with little internal structure evident; junction of epiphallus and flagellum at insertion of vas deferens, with white contents; vas deferens simple and slender, in life running beside vagina and free oviduct to join spermoviduct. Genital atrium globose, lumen almost filled with a well-developed stimulator appearing as an irregularly folded and ridged mass, flaccid rather than muscular, its contracted shape variable between individuals, but occupying most of lumen wall ( Fig. 34B View Fig ). Vagina short; gametolytic sac ovate, thin-walled, its duct short and thickerwalled; free oviduct greatly hypertrophied and convoluted, pale apricot in fresh material, occupying much of body cavity posterior to buccal mass, both dorsal and ventral to gut; spermoviduct divided into distinct prostatic and oviductal portions.
SPERMATOPHORE ( Fig. 34E View Fig ). Elbowed, with an elongate capsule and a long, spirally coiled tail of ca 6 revolutions (coiled length of tail approx. 4.1 mm); proximal portion of tail (first whorl) with paired branching spines; spines asymmetric, larger on one side; spines in middle part of this whorl largest, progressively smaller distally; spines arching outward and with branches on one side only; branch tips bluntly bifid, ending in short recurved hooks; mid-region of tail (± 1.5 whorls) lacking spines; subsequent whorls (± 2.5) of tail somewhat flattened and with short stout projections on both sides, these curve over flattened area delimiting a U-shaped channel that spirals along tail; tip of tail (± one whorl) without spines.
Distribution ( Fig. 15 View Fig )
A relatively narrow-range endemic, known only from the lower catchments of the Mbhashe and Kei Rivers (formerly known as Fingoland) and southward to the Buffalo River valley near East London; from 200 m to 835 m above sea level.
Habitat
Mostly known from Eastern Valley Bushveld ( Mucina & Rutherford 2006); living amongst the green leaf-bases of candelabra aloes.
Remarks
With its rather globose profile, the shell of Sheldonia fingolandensis sp. nov. resembles that of S. trotteriana (Benson, 1848) , but it is less elevated, has a more strongly patterned shell and more rapidly expanding whorls. Furthermore, whereas S. fingolandensis sp. nov. is arboreal, S. trotteriana is evidently exclusively ground-dwelling. The aloe-dwelling habit of the new species is shared with S. aloicola (Melvill & Ponsonby, 1890) , but that species has a more depressed, more translucent shell of a more uniform colour. It seems likely, however, that the two are closely related, as the morphology of their distal genitalia is similar. S. fingolandensis sp. nov. may well have originated as a north-eastern isolate of the more southerly and more widespread S. aloicola . The shell of S. natalensis (Pfeiffer, 1846) may also have a colour pattern of narrow axial stripes, but it is more globose, has a smaller protoconch (diameter 1.7–1.8 mm) and never attains such a large adult size (diameter <16.5 mm)
Conservation
The observations made above for Kerkophorus terrestris sp. nov. are also applicable to Sheldonia fingolandensis sp. nov.




No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |