
Riama yumborum, Aguirre-Peñafiel, Vanessa, Torres-Carvajal, Omar, Sales Nunes, Pedro M., Peck, Mika R. & Maddock, Simon T., 2014
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3866.2.4 |
|
publication LSID |
lsid:zoobank.org:pub:1688B0D1-5304-4728-8215-66B01A6C5C0D |
|
DOI |
https://doi.org/10.5281/zenodo.6133184 |
|
persistent identifier |
https://treatment.plazi.org/id/039F87A3-FFB1-E44F-FF65-DB9DE40D8242 |
|
treatment provided by |
Plazi |
|
scientific name |
Riama yumborum |
| status |
sp. nov. |
Riama yumborum sp. nov.
Proposed common name in English: Yumbo riamas
Proposed common name in Spanish: palos de los Yumbos
Holotype. QCAZ 10827 ( Figs. 1 View FIGURE 1 , 2 View FIGURE 2 , 3 View FIGURE 3 ), an adult male from Nanegal, Santa Lucía Cloud Forest Reserve, 0.11778° N; 78.607555° W (DD, WGS84), 1580–1591 m, Provincia Pichincha, Ecuador, collected on 9 March 2010 by B. Tolhurst, P. Mafla-Endara, S. Ryan, and X. Cueva.
Paratypes (5). ECUADOR: Provincia Pichincha: QCAZ 10822, female, same collection data as holotype; QCAZ 11077, 11079, 11080, 11081, males from Nanegal, Santa Lucía Cloud Forest Reserve, 0.11928° N; 78.59647° W (DD), 1585 m, Provincia Pichincha, Ecuador, collected on September 2009 by S. Maddock.
Etymology. The specific epithet, a noun in the genitive plural case which translates to “of the Yumbos,” honors the Yumbo culture (800–1660 A.D.), a pre-Incan civilization that inhabited the same area where Riama yumborum was discovered ( Chávez 2007).
Diagnosis. In addition to the molecular data discussed below, Riama yumborum can be distinguished from its congeners by the combination of two characters: incomplete nasoloreal suture and a cylindrical hemipenial body with diagonally orientated flounces on its lateral aspect. In addition, males of the new species seem to have a large number (13 per leg; n = 1) of femoral pores.
Other species with an incomplete nasoloreal suture in some individuals include Riama anatoloros ( Kizirian 1996) , R. colomaromani , R. simotera , R. stigmatoral , and R. columbiana ( Andersson 1914) . From R. anatoloros (character states in parentheses), the new species can be distinguished by usually having a single, anterior superciliary (usually complete series of four superciliaries), 13 femoral pores in males (femoral pores 7–11), and by lacking both dorsolateral stripes (such stripes present) and scales between medialmost femoral pores in males (4–12 scales). From R. colomaromani (character states in parentheses), the new species differs in having 22–23 transverse rows of ventral scales (18–21 rows), 13 femoral pores in males (femoral pores 7–9), and by lacking both scales between medialmost femoral pores in males (2–4 scales) and by lacking femoral pores in females (one pore on each side). From R. simotera (character states in parentheses), the new species can be distinguished by having usually a single, anterior superciliary (complete series of 3–4 superciliaries), striated dorsal scales (dorsals smooth), 27–29 longitudinal dorsal scale rows (19–26 rows), 13 femoral pores in males (femoral pores 5–7), and by lacking scales between medialmost femoral pores in males (2–3 scales), and by lacking femoral pores in females (5–7 pores on each side). From R. stigmatoral (character states in parentheses), R. yumborum differs in having 13 femoral pores in males (femoral pores 8–11), an average SVL = 54.5 mm (79 mm), and in lacking anterior cloacal plate scales in males (0-2). From R. columbiana (character states in parentheses) the new species is different in having striated dorsal scales (dorsals with low, rounded keel), 13 femoral pores in males (femoral pores 10), and in lacking scales between medialmost femoral pores in males (5–6 scales).
Other species of Riama with 13 or more femoral pores on each side in males are R. orcesi ( Kizirian 1995) , R. unicolor , and R. luctuosa ( Peters 1862) . The new species differs from them in having an incomplete (usually one) series of superciliaries. From R. orcesi (character states in parentheses), R. yumborum can be distinguished further by having an incomplete (complete) nasoloreal suture, dorsal scales keeled (dorsal scales striated), longitudinal dorsal scale rows in males 28–29 (13–23), and lateral scale rows 2–3 (4–8); from R. unicolor (character states in parentheses) by having 28–29 (20–26) longitudinal dorsal scale rows in males; from R. luctuosa by having an incomplete nasoloreal suture (complete in R. luctuosa ).
Hemipenial morphology can be used to distinguish Riama yumborum from all other species of Riama for which hemipenial morphology is known, except R. balneator ( Kizirian 1996) , R. cashcaensis , and R. unicolor , by having the lateral aspect of the hemipenial body ornamented with columns of diagonal flounces instead of chevronshaped or transverse flounces. It differs from R. balneator in having rows of diagonal spinulate flounces (nonspinulate flounces in R. balneator ). In lacking one or more isolated transverse spinulate flounces at the base of the asulcate side it differs from R achlyens ( Uzzell 1958) , R. afrania ( Arredondo & Sánchez-Pacheco 2010) , R. crypta ( Sánchez-Pacheco et al. 2011) , R. hyposticta ( Boulenger 1902) , R. laudahnae ( Köhler & Lehr 2004) , and R. orcesi . The new species can be further distinguished from R. crypta , R. hyposticta , and R. laudahnae by lacking distal filiform appendages on the hemipenial lobes. In having a cylindrical hemipenial body, it differs from R. achlyens , R. anatoloros , R. afrania , R. cashcaensis , R. colomaromani , R. crypta , R. hyposticta , R. laudahnae , R. striata ( Peters 1862) , R. simotera , and R. unicolor , which have an elongate, globose or conical hemipenial body.
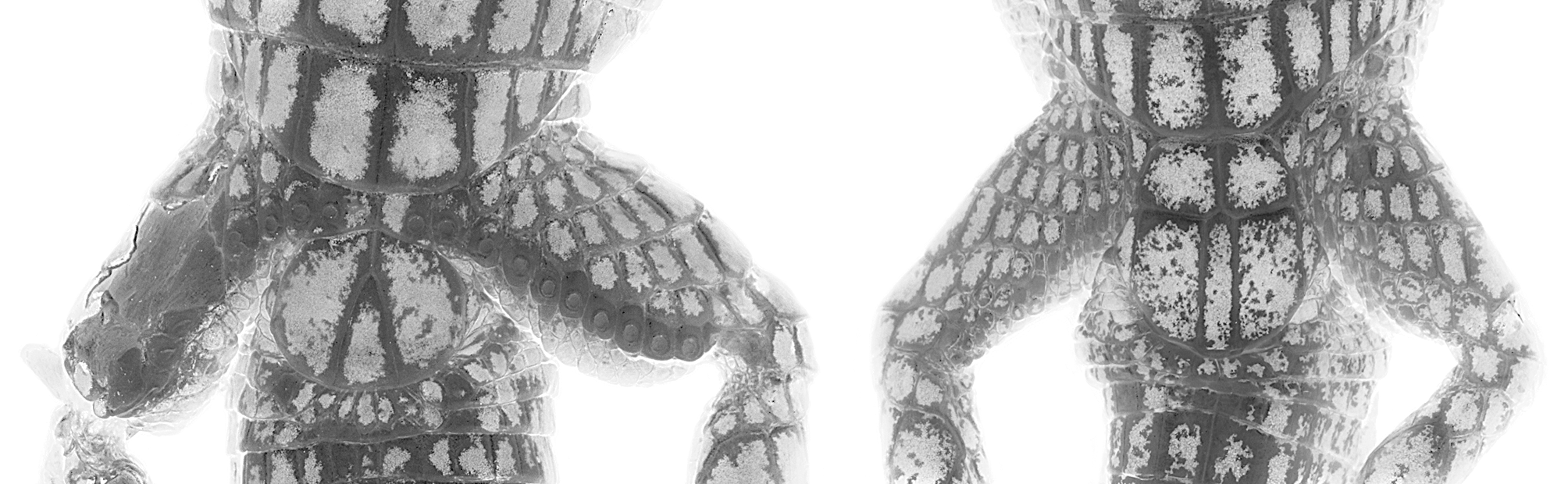
Description. Riama yumborum possesses the following characteristics: (1) SVL in adult males 51.5–59.1 mm (n = 4), in females 53.7 mm (n = 1); (2) frontonasal similar in length to frontal; (3) prefrontals absent; (4) nasoloreal suture incomplete; (5) supraoculars four; second, third and fourth in contact with ciliaries; (6) usually one superciliary (specimen QCAZ 11080 has two); (7) supralabial-subocular fusion absent; (8) postoculars three; (9) postparietals two; (10) usually two supratympanic temporals (specimen QCAZ 10827 has three on right side); (11) genials in two pairs; (12) dorsal scales rectangular, juxtaposed, striated; (13) nuchal scales smooth; (14) longitudinal dorsal scale rows in males 28–29; (15) transverse dorsal scale rows 36–40; (16) ventral scales smooth, in 22–23 transverse rows; (17) lateral scale rows two or three; (18) femoral pores in males 13 (n = 1, left side only), absent in females (n = 1); (19) scales between medialmost femoral pores in males absent; (20) subdigital scales on Toe I 4–5; (21) anterior cloacal plate absent in males, present and paired in females (n = 1); (22) dorsum dark brown, with some light-brown scales, or scales with light-brown margins; white lateral dots on neck and anterior portion of body; ventral scales dark brown with orange borders; caudal scales orange with brown pigmentation medially.
Hemipenial morphology. The left organ of QCAZ 11079 ( Fig. 4 View FIGURE 4 ) is about five subcaudal rows and six millimeters long. Hemipenial body is roughly cylindrical, ending in two symmetric small and partially everted lobes surrounded by folds; a shallow sulcus spermaticus, central in position, originates at the base of the organ and proceeds in a straight line towards the lobes.
The sulcus spermaticus ends with no clear division in independent branches as a single channel emarginated by lobular folds at the lobular crotch. Parallel to the sulcus spermaticus, two narrow nude areas bordered by rows of comb-like spicules that also ornament the lateral and asulcate sides of the hemipenial body are present.
The hemipenial body has two columns of 12 diagonal spinulate flounces (“comb-like flounces”) on each side. These flounces extend from the borders of each nude area contiguous to the sulcus spermaticus to the central asulcate face, with the five last flounces almost contacting the ones on the opposite side, forming a V-shaped structure at the sagittal region of the asulcate face. The proximal and distal flounces (in the sulcate and asulcates faces, respectively) are much reduced when compared with the intermediate ones, located on the sides of the organ.
Description of holotype (QCAZ 10827; Figs. 1 View FIGURE 1 , 2 View FIGURE 2 , 3 View FIGURE 3 , 5 View FIGURE 5 ). Male, SVL 56.5 mm, tail (complete) length 64.9 mm; head scales smooth; rostral wider than long, higher than adjacent supralabials, in contact posteriorly with nasal, frontonasal and anteriormost supralabials; frontonasal nearly rectangular in shape, widest posteriorly, similar in length to frontal, in contact laterally with nasals and posteriorly with frontal; prefrontals absent; frontal longer than wide, widest anteriorly, anterior suture slightly convex, lateral sutures slightly concave, posterior suture angular with point directed posteriorly, in contact with first and second supraoculars laterally, and frontoparietals posteriorly; frontoparietals pentagonal, in contact with second and third supraoculars laterally, parietals posterolaterally, and interparietal posteromedially; interparietal hexagonal, longer than wide, in contact with parietals laterally, and postparietals posteriorly; parietals hexagonal, in contact with third and fourth supraoculars anterolaterally, dorsalmost postoculars laterally, dorsalmost temporals posterolaterally, and postparietals posteriorly; postparietals two, longest laterally, in contact medially; supraoculars four, with second, third and fourth in contact with ciliaries; nasoloreal suture incomplete; superciliary single, located anteriorly and situated between nasal, frontal, first and second supraoculars, and anteriormost ciliaries, barely extending onto dorsal surface of head; palpebral disc semitransparent and divided into three large unpigmented scales; frenocular nearly trapezoidal in shape, in contact with nasal anteriorly; six circumorbitals between posteriormost supraocular and frenocular; postoculars three; temporals smooth; supratympanic temporals 2/3; supralabials five; infralabials four; mental wider than long, in contact with anteriormost infralabials laterally and postmental posteriorly; postmental roughly pentagonal, posterior suture angular, with angle directed posteriorly, in contact with first and second infralabials laterally; genials in two pairs, anteriormost pair roughly quadrangular, in contact with second and third infralabials; posterior pair polygonal, in contact with third and fourth infralabials; scale rows between genials and collar fold (along midventral line) eight; medialmost scales of posteriormost scale row distinctly enlarged, smooth; posteriormost gular row enfolded posteriorly, concealing three small scale rows; lateral neck scales squarish or rounded, smooth.
Dorsal scales rectangular, longer than wide, juxtaposed, striated, in 38 transverse rows on the left side (one of them incomplete) and 39 on the right one (two of them incomplete); longitudinal dorsal scale rows at fifth transverse ventral scale row 21, at 10th transverse ventral scale row 27, at 15th transverse ventral scale row 28; lateral scale rows at fifth transverse ventral scale row 7, at 10th transverse ventral scale row three, at 15th transverse ventral scale row three; lateral scales on body near insertion of forelimb small to granular; ventral scales smooth; complete transverse ventral scale rows 22; longitudinal ventral scale rows at midbody 12; anterior cloacal plate scales absent; posterior cloacal plate scales five; scales on tail rectangular and juxtaposed; midventral subcaudals smooth, wider than adjacent scales, nearly squared; femoral pores on left hind limb 13 (right limb damaged); scales between medialmost femoral pores absent. In general, limb squamation as described for congeners (e.g., Kizirian 1996).
Coloration of holotype ( Figs. 1 View FIGURE 1 , 2 View FIGURE 2 , 5 View FIGURE 5 ). In preservative (70% ethanol), dorsal ground color dark brown with scattered light brown irregular marks, more abundant on dorsal aspect of head; lateral aspect of neck and body (anteriorly) with scattered white dots; ventral scales of gular region, body, and limbs dark brown with cream borders that form a reticulate pattern; the dark pigment becomes scarce on the ventral aspect of head and tail.
Variation. The paratypes consist of a female (SVL = 53.7 mm) and four males (SVL = 31.5–59.1 mm). The paratypes are similar to the holotype with the following noteworthy exceptions. All paratypes have two supratympanic temporals on both sides (three on right side in holotype); QCAZ 10822, 11079 and 10827 have a small scale between the superciliary and adjacent ciliaries (small scale on left side only in holotype); QCAZ 11080, 11081 have two postocular on the right and left sides, respectively (three postoculars in holotype). The only female examined (QCAZ 10822) has a paired anterior cloacal plate (absent in males). In addition, QCAZ 11080 differs from all other type specimens in having a second, small superciliary (similar in size to adjacent ciliaries) in contact with the fourth supraocular on the right side, and third and fourth supraoculars on the left side.
Coloration in life of female paratype QCAZ 10822 ( Fig. 6 View FIGURE 6 ) was similar to the coloration in preservative of the holotype described above. The main difference was that in this paratype the scales of the gular region and ventral aspect of body and limbs were bordered with orange, and the tail was bright orange ventrally, with brown irregular marks.
Distribution. Riama yumborum inhabits high-elevation cloudforests on the Pacific slopes of the Andes in northwestern Ecuador ( Fig. 7 View FIGURE 7 ). It is known from Lower Montane Evergreen Forest of the Western Andes ( Iglesias et al. 2013) at elevations of 1580–1591 m in the province of Pichincha. This species occurs in sympatry with R. oculata ( O’Shaughnessy 1879) , whereas R. unicolor occurs allopatrically at higher elevations (> 1775 m) ( Maddock et al. 2011) in the same region. Other species of small ground lizards collected in the same area include the sphaerodactylid gecko Lepidoblepharis conolepis Avila-Pires 2001 , and the gymnophthalmids Alopoglossus viridiceps Torres-Carvajal & Lobos 2014 , Cercosaura vertebralis O’Shaugnessy 1879 and Echinosaura brachycephala Köhler et al. 2004 .
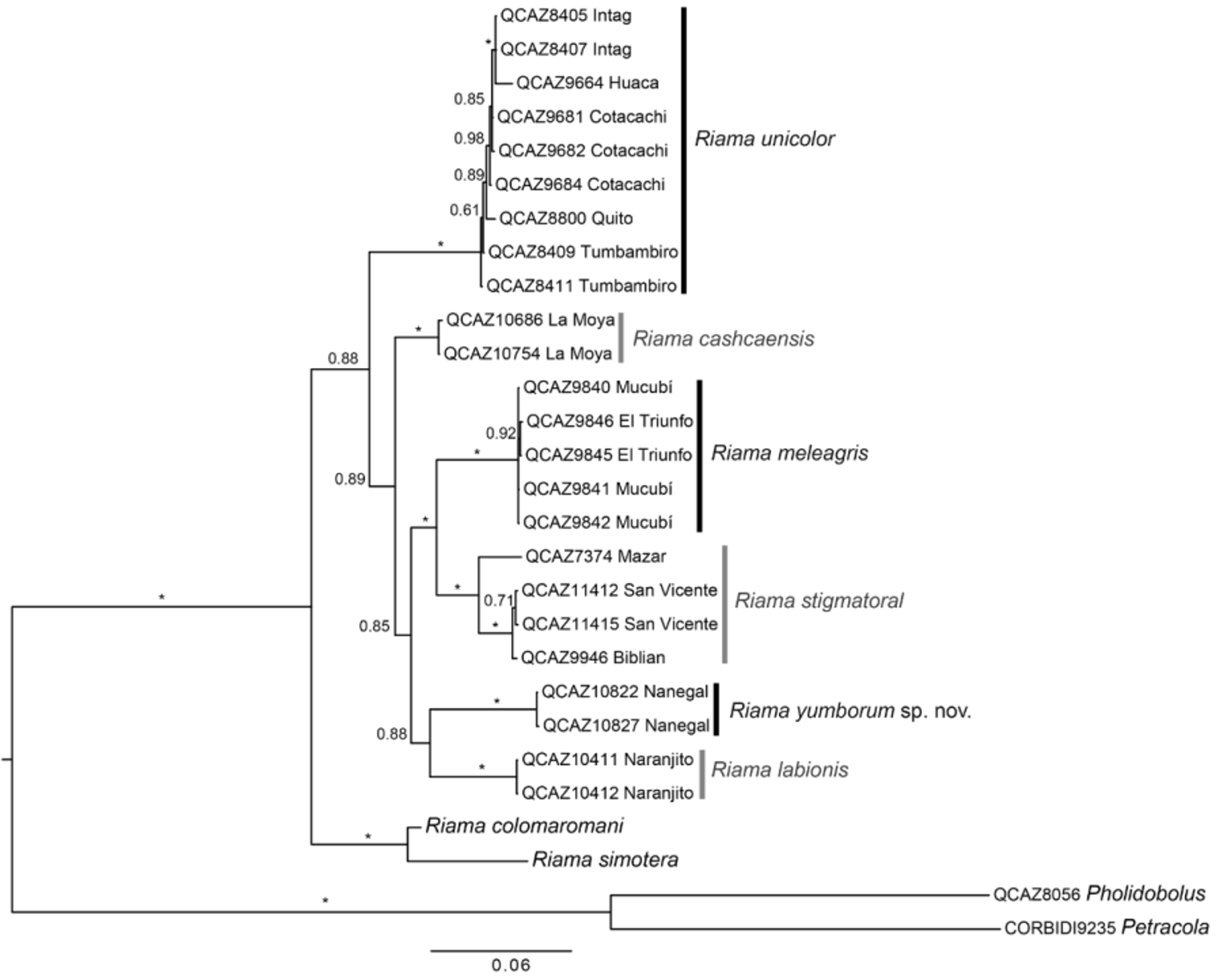
Phylogenetic relationships. Selected models of evolution of gene regions used in phylogenetic analyses were TIM2+I+G for 12S and 16S; GTR+I+G, TIM3+I+G, TIM2+I+G for first, second, and third codon positions of ND4, respectively; and HKY, TPM1uf+I, TPM3uf for first, second, and third codon positions of c-mos, respectively. The inferred phylogenetic tree ( Fig. 8 View FIGURE 8 ) is generally concordant with the phylogenetic tree of Castoe et al. (2004), although their analyses did not include Riama yumborum , R. labionis , R. meleagris , and R. stigmatoral . Our tree strongly supports (PP = 1) the monophyly of Riama and a clade consisting of R. yumborum , R. labionis , R. stigmatoral , R. meleagris , R. cashcaensis , and R. unicolor . Relationships among these inner taxa are less supported (PP <0.89), except the sister species relationship of R. meleagris and R. stigmatoral , and R. colomaromani and R. simotera (PP = 1). The sister taxon of R. yumborum is R. labionis (PP = 0.88); and both are sister (PP = 0.85) to the R. meleagris-stigmatoral clade ( Fig. 8 View FIGURE 8 ).






| QCAZ |
Museo de Zoologia, Pontificia Universidad Catolica del Ecuador |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.