
Syllis patersoni, Soto & Martín & Lucas, 2020
|
publication ID |
https://doi.org/10.11646/zootaxa.4878.1.6 |
|
publication LSID |
lsid:zoobank.org:pub:ED7F02DC-27B8-4BB8-A4C7-EC1995D5C473 |
|
DOI |
https://doi.org/10.5281/zenodo.4425543 |
|
persistent identifier |
https://treatment.plazi.org/id/039EEC36-FFC4-9B53-399A-FC38FAD4FE84 |
|
treatment provided by |
Plazi |
|
scientific name |
Syllis patersoni |
| status |
sp. nov. |
Syllis patersoni View in CoL new species
Figure 7 View FIGURE 7
Material examined. St. 41. Holotype ( MNCN 16.01 About MNCN /18728) and two Paratypes ( MNCN 16.01 About MNCN /18764). Date of collection: 19 October 2009 (St. 41). Coordinates: St. 41 50°16’37” S 74°53’21’’W GoogleMaps .
Description. Body elongated, slender ( Fig. 7A View FIGURE 7 ), without color markings; holotype complete, 19 mm long, 0.5 mm wide, 132 chaetigers. Prostomium almost circular, with two pairs of small eyes in open trapezoidal arrangement; median antenna slightly longer than prostomium and palps together, inserted in front of posterior eyes, with about 19 articles; lateral antennae inserted in front of anterior eyes, shorter than median antenna, with 18 articles. Peristomium distinctly shorter than subsequent segments ( Fig. 7A View FIGURE 7 ); dorsal tentacular cirri longer than median antenna, with 25 articles; ventral tentacular cirri shorter than dorsal ones, with 13 articles. Dorsal cirri similar in length to body width, longer on anterior segments, slender, elongated, with numerous spiralized glands inside articles ( Fig. 7A, B View FIGURE 7 ); anterior dorsal cirri somewhat longer than remaining, with about 27-14-20-27-17 articles on first five chaetigers; dorsal cirri from proventricular segments alternating long cirri, with 29 articles, and short ones, with 19 articles ( Fig. 7B View FIGURE 7 ); articles plenty of distinct refringent glands ( Fig. 7A, B View FIGURE 7 ). Parapodia conical; ventral cirri digitiform. Compound chaetae composed by few short, spiniger-like chaetae ( Fig. 7C, E, G View FIGURE 7 ) and several heterogomph falcigers with elongated, slender, bidentate blades, proximal teeth slightly smaller than distal ones, and short to moderate spines on margin, with dorso-ventral gradation in length ( Fig. 7D, F, H View FIGURE 7 ), similar throughout. Anterior parapodia each with 2–3 short spiniger-like chaetae, blades 69 µm ( Fig. 7C View FIGURE 7 ) and 11–12 falcigers with dorso-ventral gradation, blades 36 µm above, 22 µm below ( Fig. 7D View FIGURE 7 ); midbody parapodia each with two spiniger-like chaetae, blades 75– 71 µm long ( Fig. 7E View FIGURE 7 ) and 6–7 falcigers, blades 35 µm above, 20 µm below ( Fig. 7F View FIGURE 7 ); posterior parapodia each with one spiniger-like, shorter than those of midbody and anterior segments, blades 57 µm long ( Fig. 7G View FIGURE 7 ), and 5–6 falcigers, blades 29 µm above, 16 µm below ( Fig. 7H View FIGURE 7 ). Solitary dorsal capillary chaetae on posterior parapodia, slender bidentate, with minute subdistal serrations ( Fig. 7I View FIGURE 7 ); ventral simple capillary chaetae on far posterior segments, more distinctly bidentate than dorsal one ( Fig. 7J View FIGURE 7 ). Anterior parapodia with four slender, pointed aciculae ( Fig. 7K View FIGURE 7 ), three on midbody parapodia, distinctly thicker ( Fig. 7L View FIGURE 7 ), and single in posterior parapodia, acuminate ( Fig. 7M View FIGURE 7 ). Pharynx long and slender, through more than 12 segments (everted); pharyngeal tooth conical, on anterior margin ( Fig. 7A View FIGURE 7 ). Proventricle slender and long, shorter than pharynx, through eight segments, with 42 muscle cell rows, without midline ( Fig. 7A View FIGURE 7 ). Pygidium small, with two long anal cirri, with 25 articles.
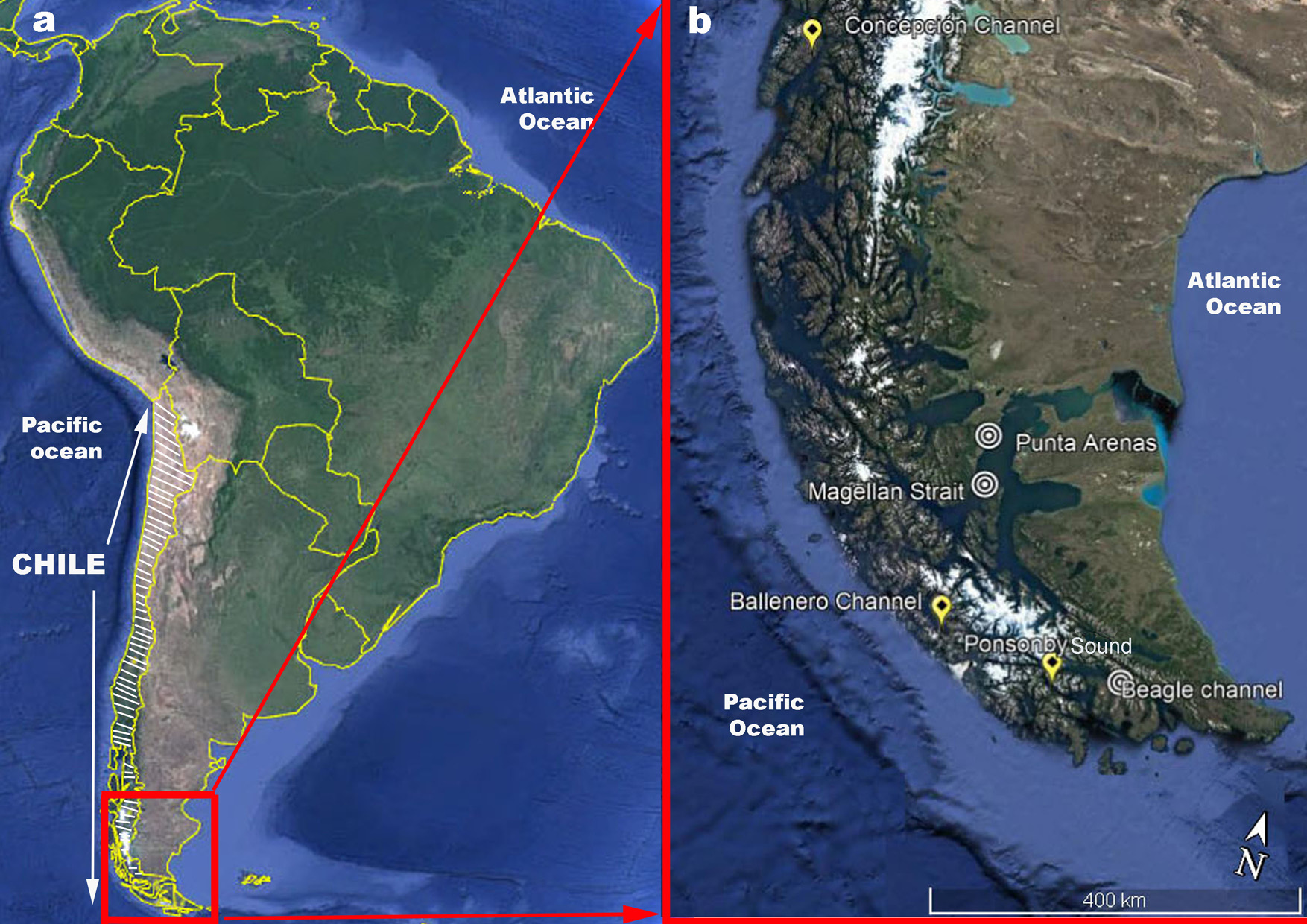
Distribution. Only known from Chilean Patagonia in Concepción channel, Drumond Hay island ( Fig. 1 View FIGURE 1 ).
Habitat. From boulders, sediment bottoms and associated to M. pyrifera holdfasts in fjords, islands and channels from Chilean Patagonia. Salinity: from 30.2 PSU, temperature: 8.9°C. Shallow subtidal at 30 m depth. This species was not found inside tubes of Chaetopterus cf. variopedatus .
Type locality. Concepción channel, Drumond Hay island, southern Chile (Patagonia) .
Remarks. Syllis patersoni , n. sp. is characterized by its elongated, slender body, long pharynx and proventricle, acuminate posterior acicula, and two types of compound chaetae on each parapodium, relatively short spiniger-like chaetae and falcigers, both bidentate, with proximal teeth well-marked, and distal teeth slightly rounded, and short to moderate straight spines on margin.
Syllis alosae San Martín, 1992 , from Cuba, also has similar falcigers, but the spiniger-like are distally broad, shows a marked color pattern, the dorsal cirri are shorter, and the posterior acicula are straight, protruding out from the parapodial lobes ( San Martín 1992). Syllis maryae San Martín, 1992 , from Cuba and Brazil, has similar body, dorsal cirri, pharynx and proventricle, but the spiniger-like chaetae are proportionally longer, in relation to the falcigers, than those of Syllis patersoni , n. sp. The posterior acicula are distally bent at a right angle, and the dorsal simple chaetae are truncate; furthermore, some specimens show some dorsal glands on some post-proventricular segments ( San Martín 1992; Nogueira & San Martín 2002). Similar differences are shared with Syllis edensis ( Hartmann-Schröder, 1989) from Australia ( Hartmann-Schröder 1989; Álvarez-Campos et al. 2015b). Syllis bouvieri Gravier, 1900 , from the Red Sea, also has similar type of compound chaetae but the longer ones are not so long in proportion with the falcigers, the proximal and distal teeth are more separated, with a curved arc in between, the spines on margin of blades are longer, the dorsal cirri are longer, and the proventricle is shorter and wider ( Gravier 1900; Lucas et al. 2020). Syllis hyllebergi ( Licher 1999) , from the Eastern Mediterranean, Red Sea and Brazil, has shorter dorsal cirri, shorter proventricle, and the falcigers are more markedly bidentate, with longer spines on margin ( Licher 1999; Nogueira & San Martín 2002). Several other species also have spiniger-like and falciger chaetae, but the spiniger-like chaetae are much more longer and slender, with filiform blades, in proportion with the falcigers.
Syllis beneliahuae ( Campoy & Alquézar 1982) , widely distributed in tropical and temperate seas, is also a similar species, with slender body, and relatively short spiniger-like chaetae, acuminate posterior acicula and relatively long dorsal cirri, with articles with plenty of refringent glands; however that species has longer spiniger-like chaetae (blades more than twice as long as the longer falciger) and shorter proventricle ( Campoy & Alquézar 1982; Campoy 1982; San Martín 2003).
Syllis augeneri Haswell, 1920 , from Australia and Indonesia, also has acuminate posterior acicula and short spiniger-like chaetae, but the dorsal cirri are much shorter and proportionally thicker, and the spiniger-like chaetae are much shorter than those of S. patersoni n. sp. ( Haswell 1920; Aguado et al. 2008).
Etymology. This species is named in honor of Dr. Gordon LJ Paterson, British polychaetologist currently retired.
Discussion and conclusion s
It is well-known that some invertebrates and biological substrates are important habitats for polychaetes ( Alvarez-Campos & Verdes 2017; Martin & Britayev 1998). Syllis species presented on this work were collected from biological substrates such as Macrocystis pyrifera (Linnaeus) C. Agardh, 1820 kelp holdfasts and Chaetopterus cf. variopedatus ( Renier, 1804) tubes ( Chaetopteridae ) as well as from pebbles and sandy bottoms. In this research three new Syllis species were described; two of them were found associated to C. cf. variopedatus and all three species were found living on Macrocystis pyrifera holdfasts. Chaetopterus tubes are a new habitat for Syllis possibly as an adaptation to the lack of other suitable habitats due to the presence of extreme environmental conditions and the influence of ice-fields present in the fjords and channels of the Chilean Patagonia. Chaetopterus tubes have demonstrated to be a suitable habitat for several syllid species previously recorded at Chilean Patagonia by Soto & San Martin (2017 a,b). These authors have already found twelve species of syllids hence with the current work the total number of syllid species increases to 15. High abundance and diversity of syllids in association with biological substrates have been previously reported ( Hernández et al. 2001; Álvarez-Campos & Verdes 2017; Soto & San Martin 2017a, b). Chaetopterid tubes clearly favour symbiotic associations offering a good habitat for diverse species, due to well-protected shelter and continuous water flow, likely bringing easily accessible food particles ( Britayev and Martin 2019). The great majority of tubes were occupied by one Chaetopterus species although Syllis species were also recorded inside empty tubes and even between the layers that form these tubes. Similarly, M. pyrifera holdfasts also allow that several syllid species may inhabit them. Nine species have been recorded by Soto and San Martin (2017a, b) for Chilean Patagonia and with this work the total number of syllids increase to 12. Syllis seems to be a common inhabitant on this type of habitat because Alvarez-Campos and Verdes (2017) recorded nine species of this genus and total number of twelve species of Syllidae inhabiting holdfasts of the brown algae Lessonia spicata in Central Chile. These authors highlighted the importance of kelp holdfasts as hotspots for syllid diversity, understanding that kelp-like species play an important ecological role providing food, mechanical shelter against wave impact, refuge from predators, secure spawning areas and nursery grounds ( Pabis and Sicinski 2010).

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Syllinae |
|
Genus |