
Vignadula kuraburiensis, Tan & Tan & Sanpanich & Duangdee & Ambarwati, 2022
|
publication ID |
https://doi.org/ 10.1093/zoolinnean/zlac031 |
|
publication LSID |
lsid:zoobank.org:pub:968C576E-4747-4586-9D48-C24C92741905 |
|
DOI |
https://doi.org/10.5281/zenodo.7036973 |
|
persistent identifier |
https://treatment.plazi.org/id/F15E6689-04B3-43C6-B44E-7D98457FFB98 |
|
taxon LSID |
lsid:zoobank.org:act:F15E6689-04B3-43C6-B44E-7D98457FFB98 |
|
treatment provided by |
Plazi |
|
scientific name |
Vignadula kuraburiensis |
| status |
sp. nov. |
VIGNADULA KURABURIENSIS View in CoL SP. NOV.
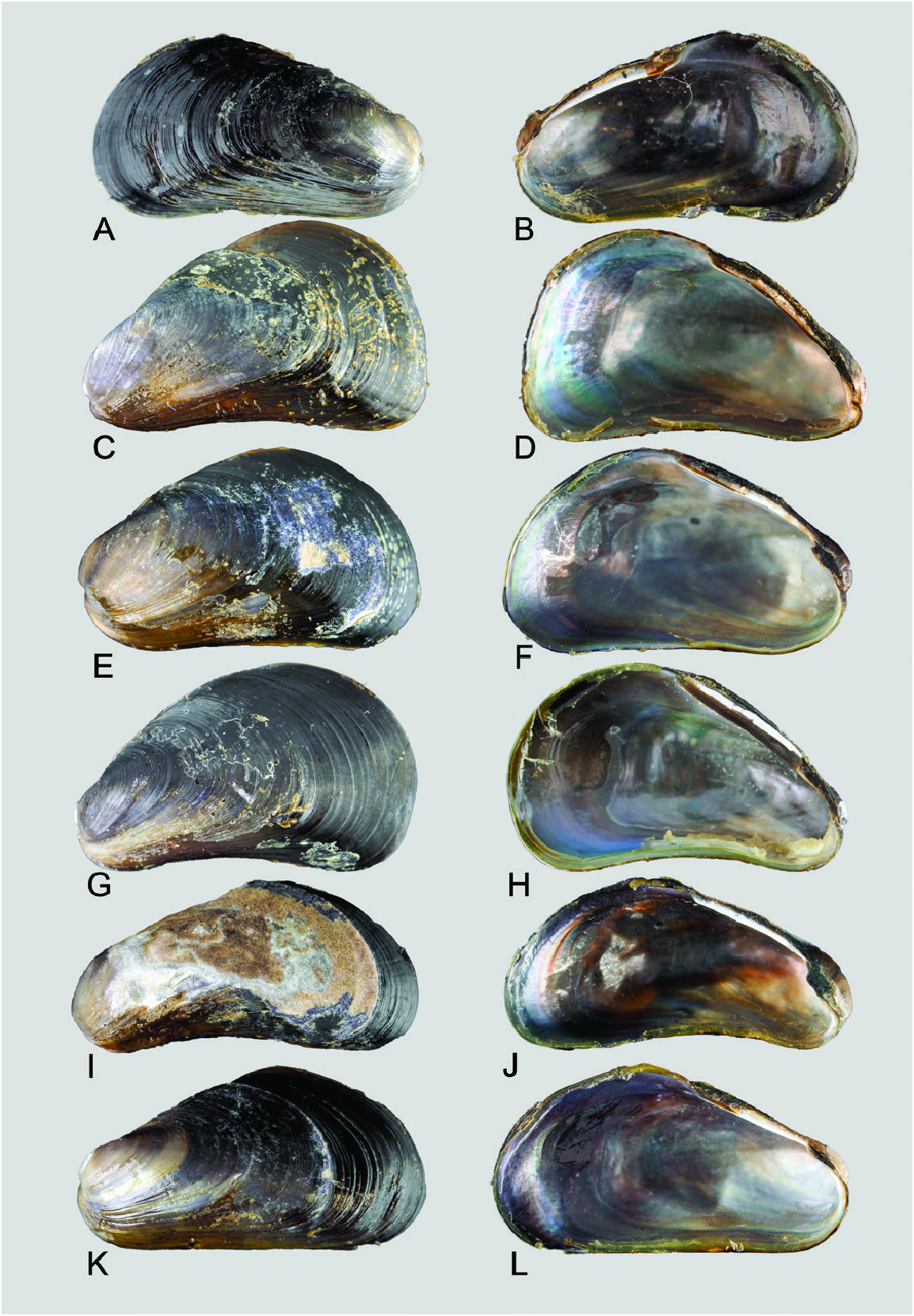
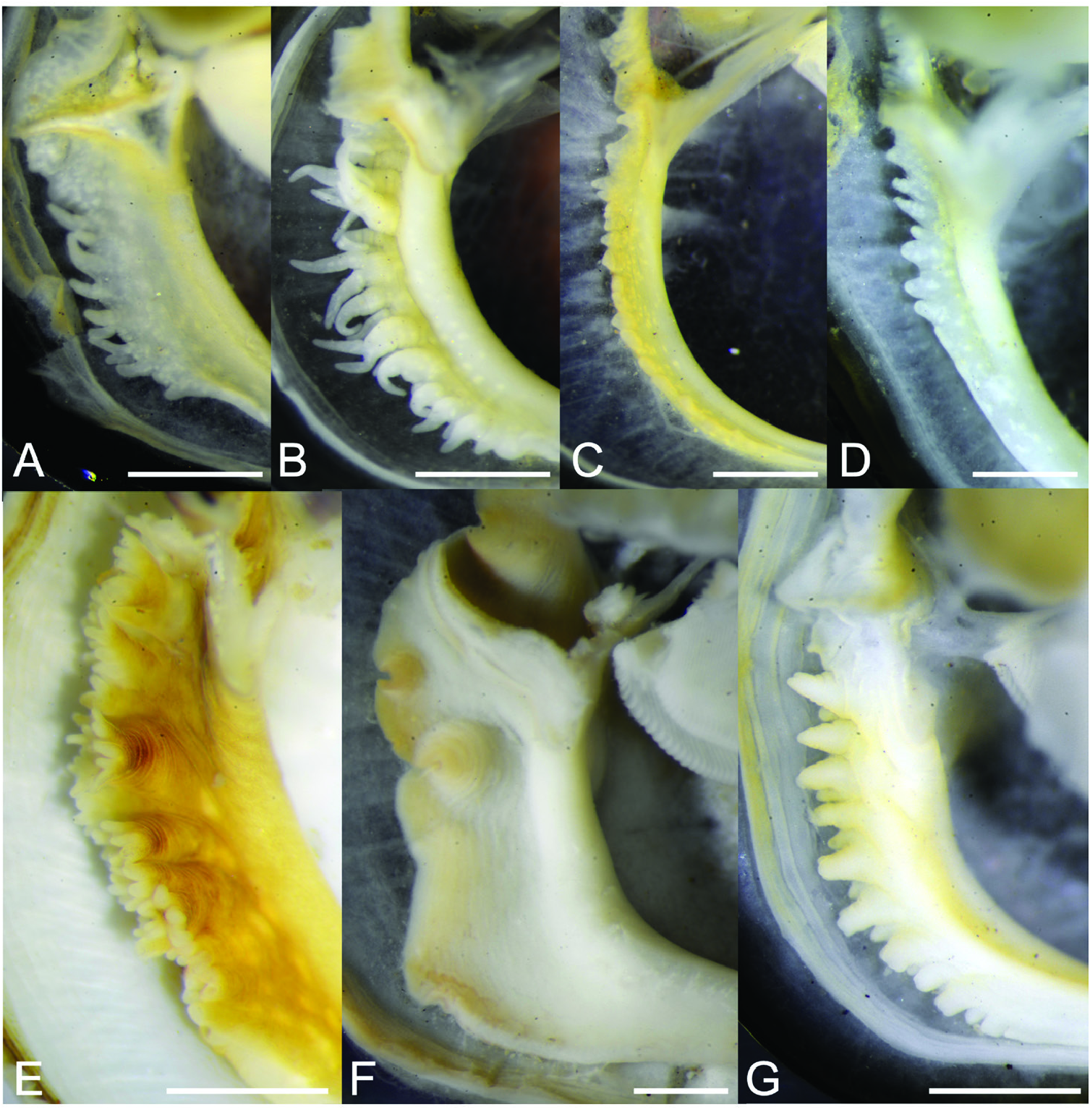
( FIGS 1–3 View Figure 1 View Figure 2 View Figure 3 , 7 View Figure 7 , 11 View Figure 11 , 12C, D View Figure 12 , 13 View Figure 13 )
ZooBank registration.: urn: lsid: zoobank. org:act: F15E6689-04B3-43C6-B44E-7D98457FFB98
Diagnosis: Light brown to dark purple mytiliform shells with terminal umbones; anterior half of ventral region often golden yellow; animal without plicate gland; mantle edge at posterior region of animal bears ≤ 18 short, simple guard papillae.
Etymology: Named after the district of Kuraburi, Phang-nga Province in south-west Thailand, where the species was first collected. The gender of the genus Vignadula ( Kuroda & Habe, 1971) was not given explicitly in the original description, but is here assumed to be female because the species name of the type species was changed from atratus to atrata ( Kuroda et al., 1971: 348, 549).
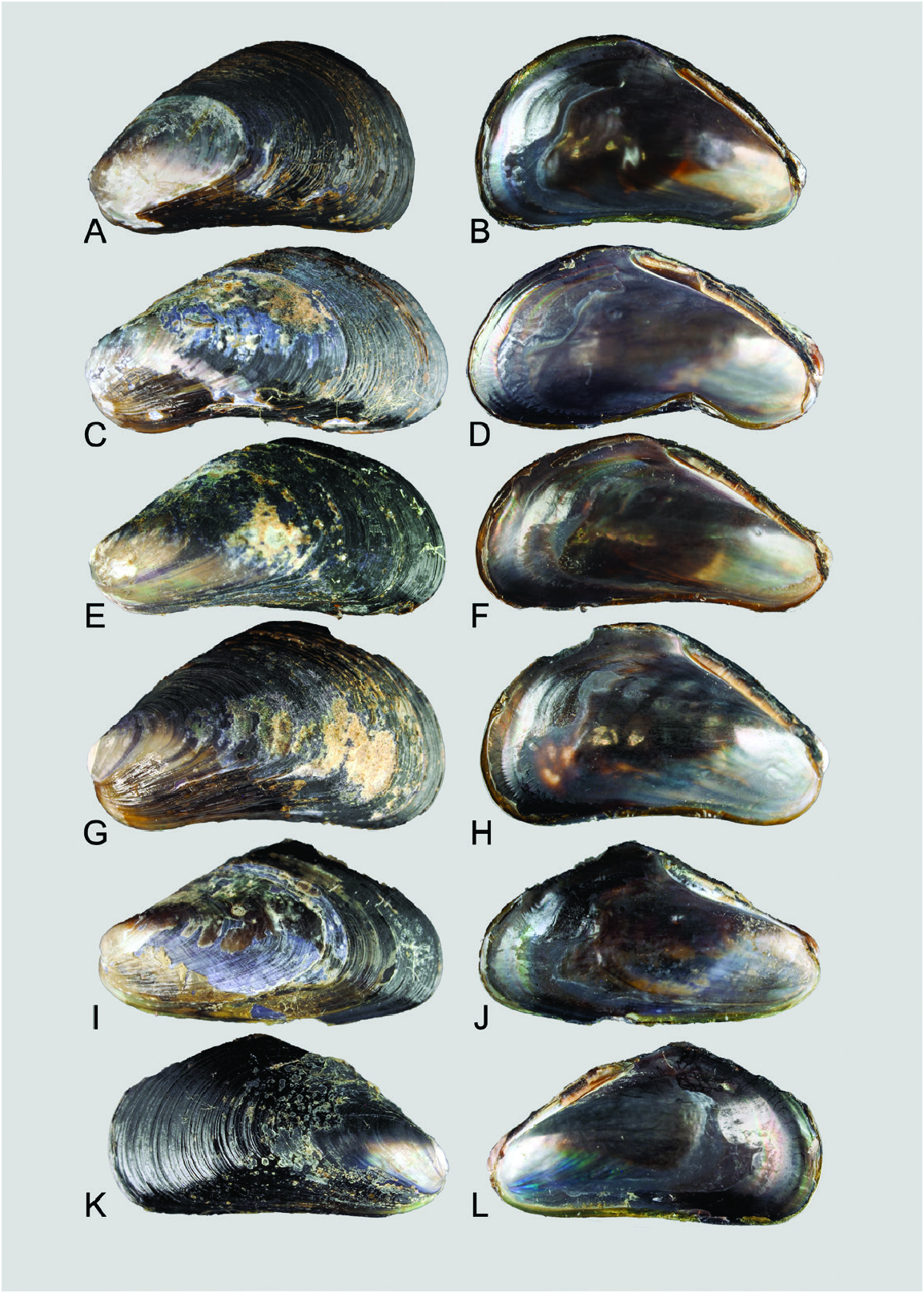
Material examined: Holotype: PMBC 25290 View Materials , Ta Bon Bang Wan jetty south of the mouth of Baan Tang La Ong, Kuraburi District, Phang-nga Province, Thailand; SL = 12.8 mm (sequenced, XTSW1 ; Fig. 7A, B View Figure 7 ) . Paratypes: PMBC 25291 View Materials , SL = 13.2 mm ( Fig. 7C, D View Figure 7 ) ; ZRC.MOL.24068, SL = 12.3 mm (sequenced, XTSW3 ; Fig. 7E, F View Figure 7 ) ; PMBC 25291 View Materials (sequenced, XTSW4 ) ; ZRC. MOL.24069, 24083 (intact individuals); Bang Ben Bay , Laem Son National Park, Ranong Province, Thailand , ZRC.MOL.24070 (sequenced, XTSW5 ), 24084 (intact individuals) .
Shell: Length ≤ 15 mm, thin, outline mytiliform; anterior region narrow, umbones terminal, barely separated from anteroventral extension; posterior region expanded, flat. Surface light brown to dark purple over dorsal region and posterior two-thirds of length of valves, anteroventral region golden yellow. Valve surfaces with fine, closely set commarginal lines that may be outlined in a darker colour. ‘Special byssal secretions’ (sensu Ockelmann, 1983; i.e. commarginal thickenings or serrations, presumably secreted by the foot) may be present. Umbones and anteroventral margin are in line. Prodissoconch II diameter ~210– 230 µm. Shell interior iridescent, usually reflecting the colour of the exterior; margins devoid of teeth. Ligament narrow, smooth without teeth; resilial pits absent.
Shell microstructure: Shell generally comprises two layers below the periostracum: a thin prismatic layer, under which a thick sheet nacreous layer is present. A thin, fibrous prismatic myostracum may be present within the sheet nacreous layer.
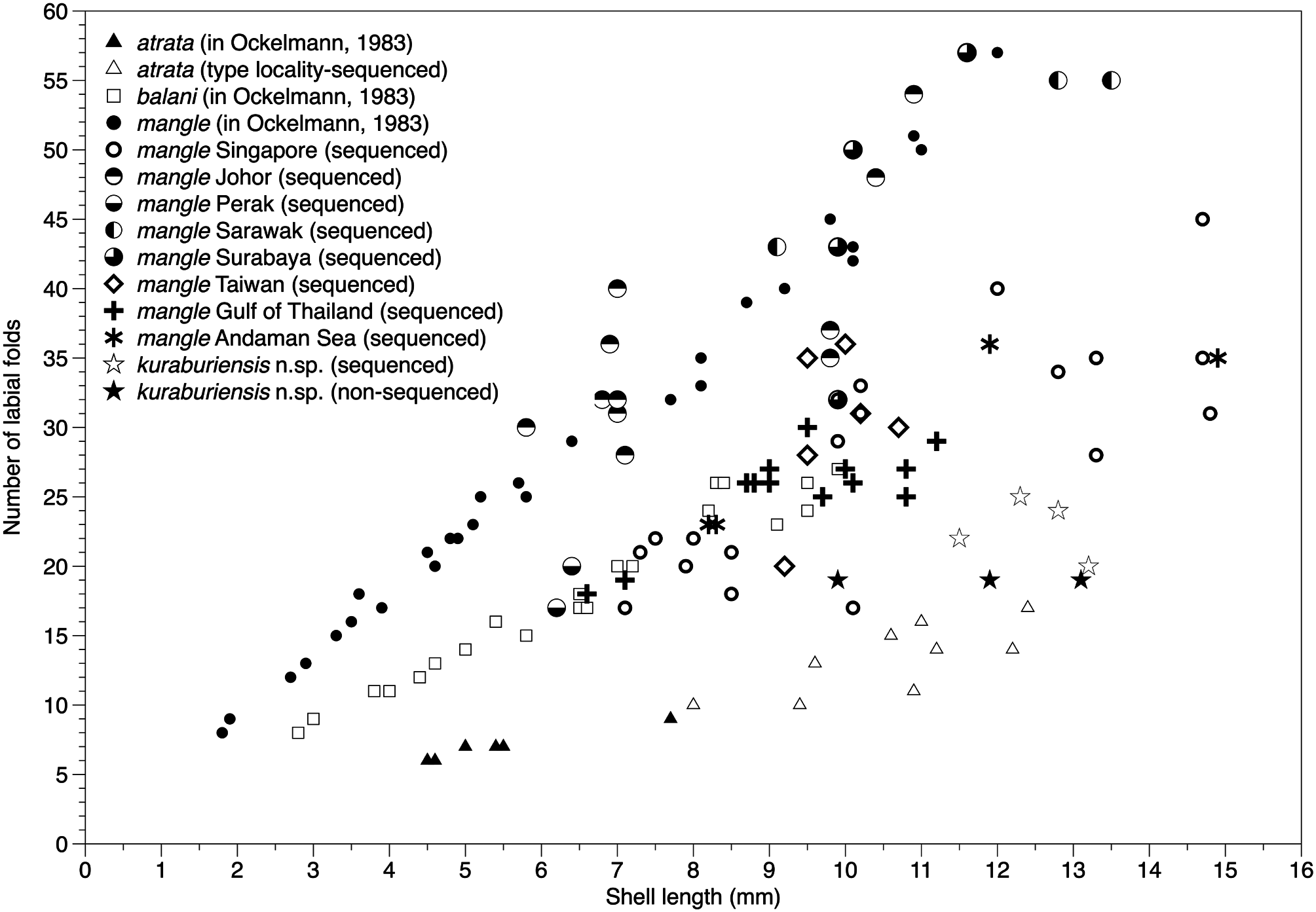
Anatomy: Up to 18 short, simple guard papillae along each side of inner mantle edge forming the inhalant aperture. Papillae translucent white, with opaque white subcutaneous pigment. Posterior byssal retractor muscles are split into two bundles (as observed on animal, although the muscle scars on the valves do not reflect this separation). Pericardium lies anterior to the posterior byssal retractor muscle complex (Category 2 of Morton, 2015a). Plicate organ absent. Foot yellowish white, vermiform. Labial palps vary in length according to shell size (see Theisen, 1982; Ockelmann, 1983), as does the number of folds (sorting ridges), which can range between 18 and 24 for SLs 10–13 mm ( Fig. 11 View Figure 11 ).
Habitat: Mid- to high intertidal, amongst oysters ( Saccostrea Dollfus & Dautzenberg, 1920 ) on concrete pilings under jetty near mangroves.
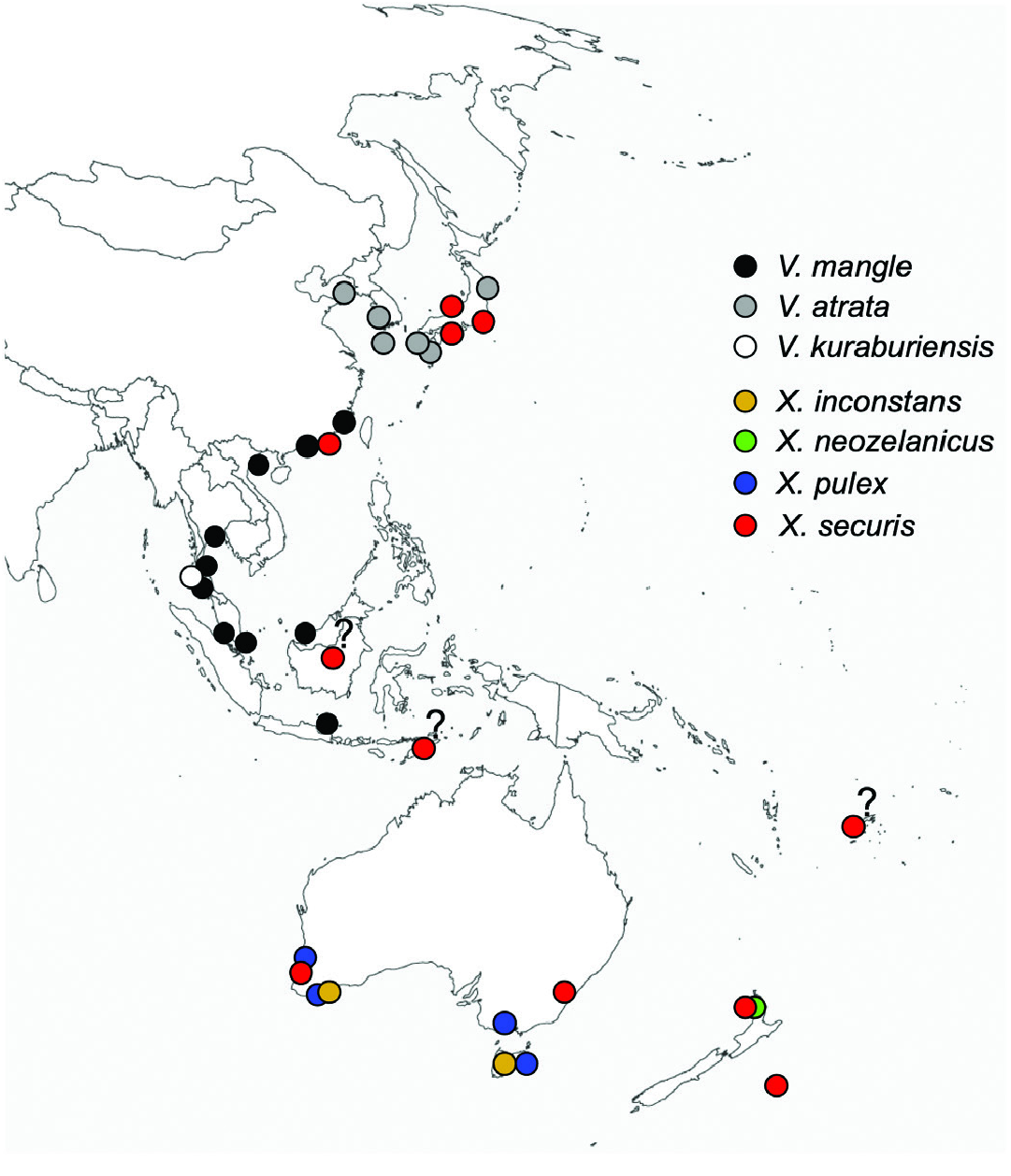
Geographical distribution ( Fig. 13 View Figure 13 ): So far, found only in Phang-nga and Ranong provinces along the Andaman Sea coast of Thailand.
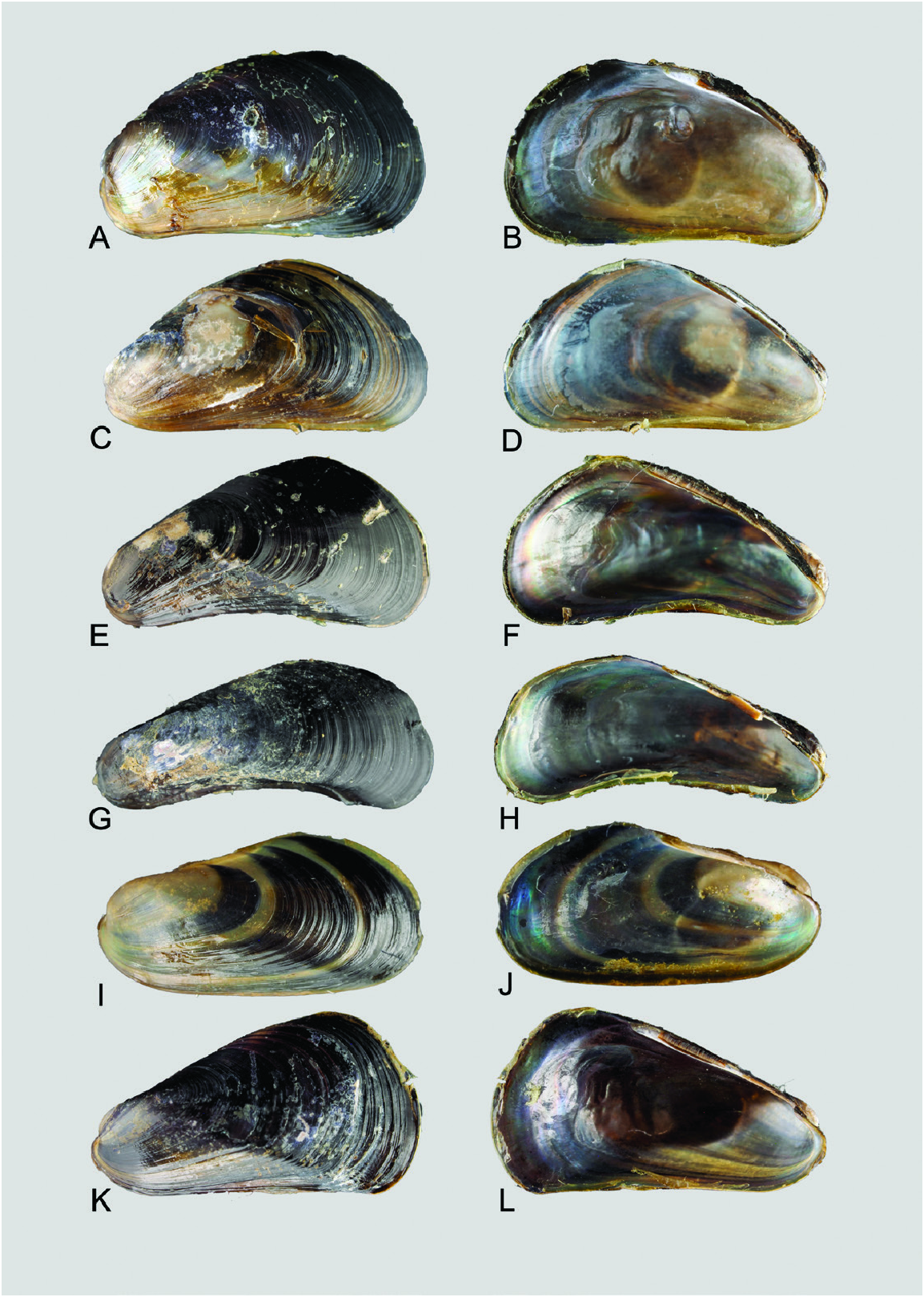
Similar species: Owing to the inherent variability of shell outline and colour in V. atrata and V. mangle (see Figs 4–6 View Figure 4 View Figure 5 View Figure 6 ), it is generally difficult to separate the new species from its congeners based solely on external appearance. However, in anatomical terms V. kuraburiensis is distinguished from V. mangle in having fewer folds on its labial palps (<20 folds in an animal ~ 12–14 mm SL; see Fig. 11 View Figure 11 ) in comparison to the more numerous (> 30 folds) labial palp folds in V. mangle ( Table 1 View Table 1 ). In addition, the absence of a plicate organ (organ of Sabatier; Thomsen et al., 2018) at the base of the ctenidium in the new species also clearly distinguishes it from V. mangle . As for V. atrata , it is anatomically similar to V. kuraburiensis (i.e. small labial palps and absence of plicate glands), and although the two species are genetically distinct and do not overlap in geographical distribution, they cannot be separated reliably using external or internal morphology.


| ZRC |
Zoological Reference Collection, National University of Singapore |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.