
Vignadula mangle Tan & Tan & Sanpanich & Duangdee & Ambarwati, 2022
|
publication ID |
https://doi.org/ 10.1093/zoolinnean/zlac031 |
|
publication LSID |
lsid:zoobank.org:pub:968C576E-4747-4586-9D48-C24C92741905 |
|
DOI |
https://doi.org/10.5281/zenodo.7043593 |
|
persistent identifier |
https://treatment.plazi.org/id/039E6F24-132C-FFEC-6553-B6DCFA4BB22E |
|
treatment provided by |
Plazi |
|
scientific name |
Vignadula mangle |
| status |
comb. nov. |
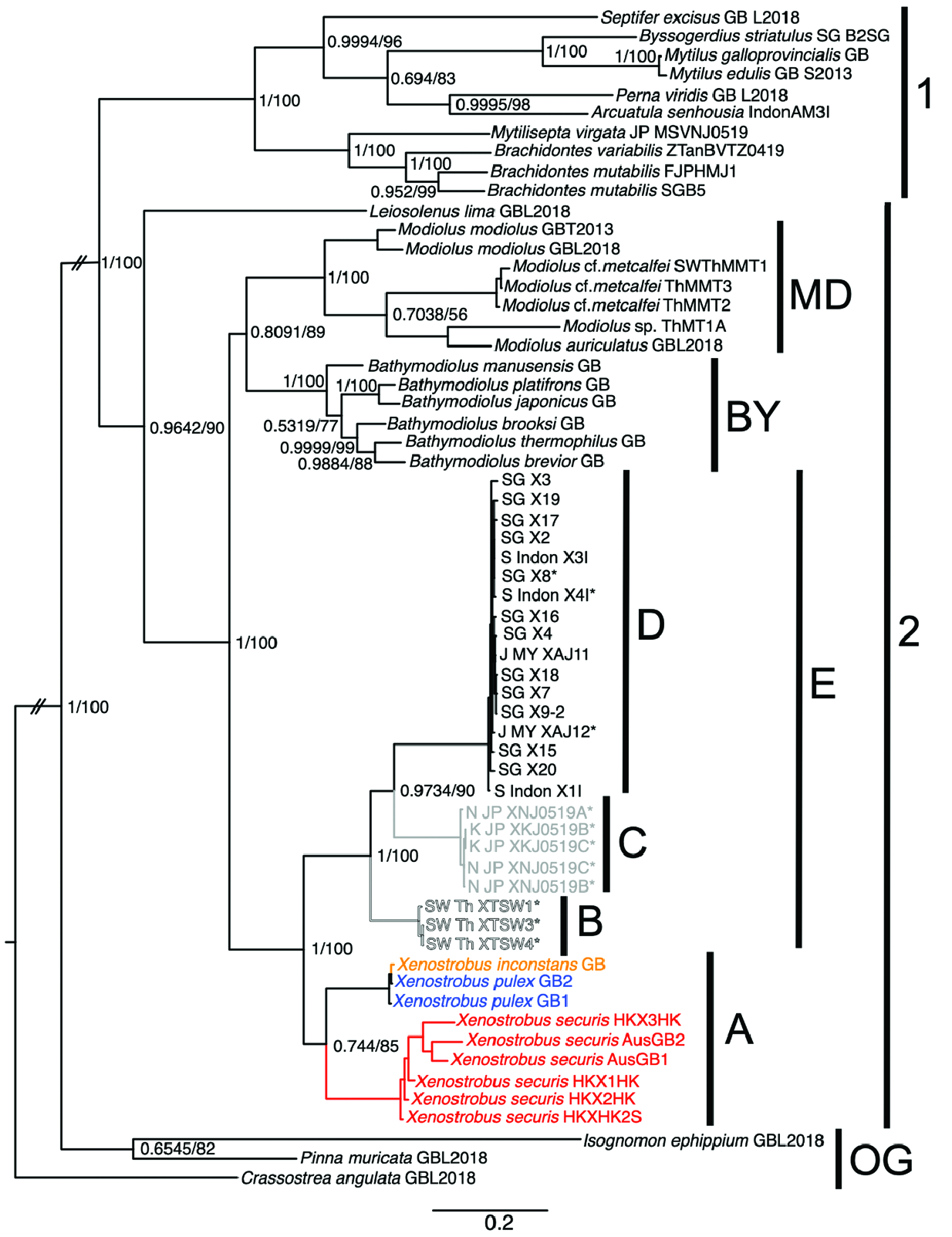
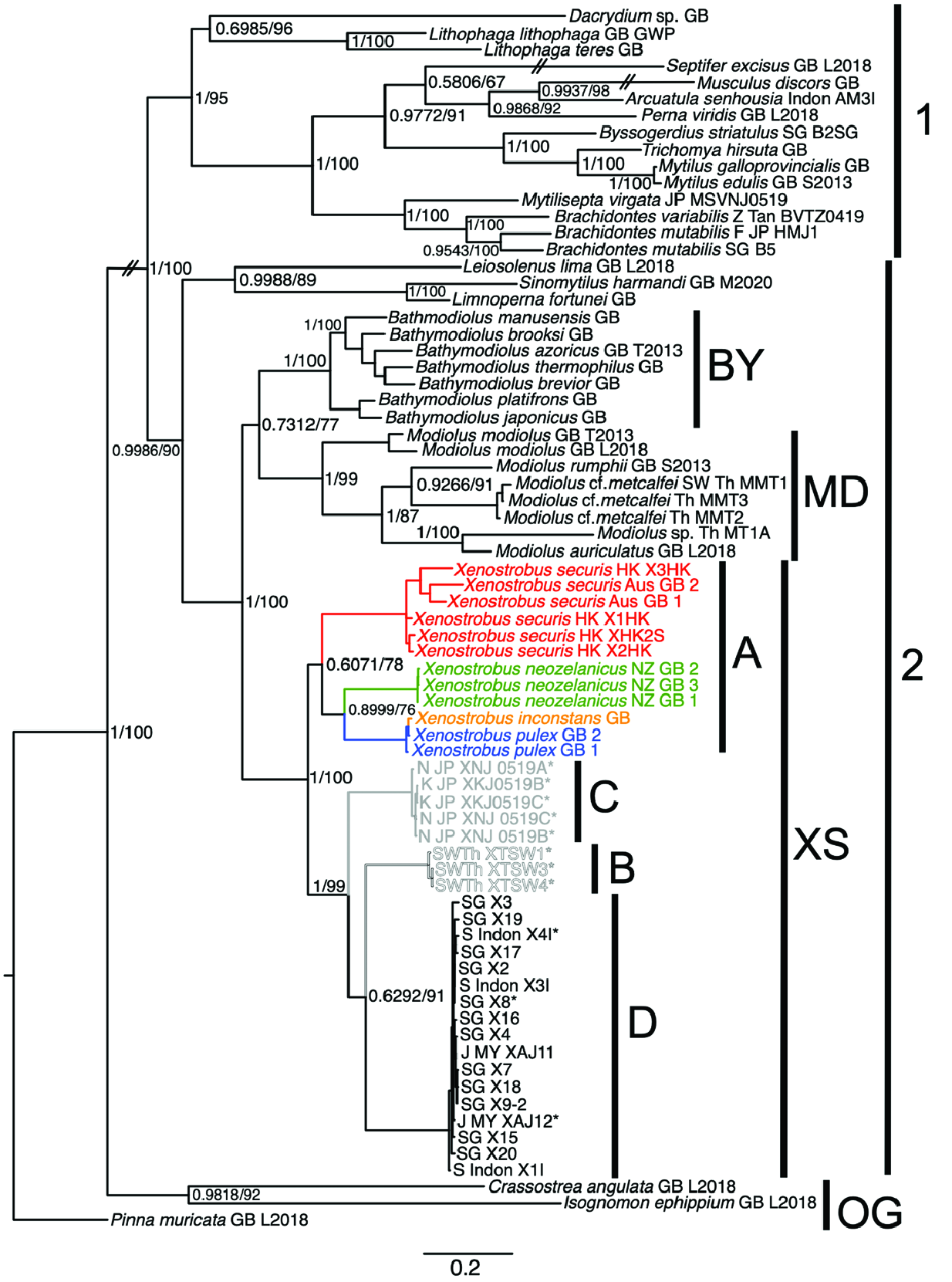
VIGNADULA MANGLE ( OCKELMANN, 1983) View in CoL COMB. NOV.
( FIGS 1–3 View Figure 1 View Figure 2 View Figure 3 , 5 View Figure 5 , 6 View Figure 6 , 9–11 View Figure 9 View Figure 10 View Figure 11 , 12B View Figure 12 , 13 View Figure 13 )
Xenostrobus mangle Ockelmann, 1983: 104–106 View in CoL , figs 32–35, 38, 40, 43, 44;
type locality: Jeram , Kuala Selangor, west coast of Malaysia. Holotype NHMD 77043 (BIV-001811) : missing; NHMD 916051 : 57 paratypes (27 intact individuals and 30 valves).
Xenostrobus balani View in CoL – Ockelmann, 1983: 107–111, figs 36, 37, 39, 41, 43–45; type locality: mangrove of Ao Nam Bor, Phuket, Thailand. Holotype missing. NHMD 915909: five paratypes; PMBC 3104: three paratypes ( Aungtonya et al., 1999).
Xenostrobus aratus (Dunker, 1856) – Nguyen, 2001: 417 (not aratus [sic]).
Xenostrobus atrata (Lischke, 1871) View in CoL – Lee & Morton, 1985: 54–55, pl. 2A (not atrata View in CoL ); – Wang, 1997 (in part): 208, fig. 87 (not atrata View in CoL ).
Xenostrobus atratus View in CoL – Bernard et al., 1993: 33–34 (in part); – Kurozumi, 2000: 429–430 and pl. 429, fig. 6 (not atratus View in CoL ?); – Kurozumi, 2017: 1173 and pl. 472, fig. 9 (not atratus View in CoL ?).
Xenostrobus sp. – Ng & Sivasothi, 1999: 119, fig.
Diagnosis: Dark purple to black mytiliform shells with a trapezoid outline and mostly subterminal umbones; anterior half of ventral region often yellow to orange crossed by dark purple commarginal lines; animal with inner and outer plicate glands; labial palps long, each palp with ≤ 60 folds; mantle edge at posterior region of animal bears ≤14 simple and/or branched guard papillae. Nomenclature: Ockelmann (1983) designated and illustrated the holotype of X. mangle but did not isolate the specimen from amongst some 27 intact individuals and 30 valves designated paratypes at NHMD. However, none of the paratype shells in NHMD matches the overall appearance and dimensions of the holotype provided by Ockelmann. In the case of X. balani , the holotype specimen is missing from the vial marked holotype in the NHMD collection (T. Schiøtte, pers. comms.). The ten separated valves (referable to five individuals) in the paratype lot at NHMD all do not match exactly the holotype dimensions given by Ockelmann (1983). Nevertheless, these irregularities do not affect the conclusions reached in this study.
Material examined: Taiwan: Husia, Kinmen (off Xiamen, China) (ZRC.MOL.24925); Vietnam: Sung Sot Cave, Ile de la Surprise off Cat Ba Island, Gulf of Tonkin
(ZRC.MOL.24926); Thailand: Andaman Sea–Pak Bara, Satun (ZRC.MOL.24927, 24928; PMBC 25331); Ko Talibong, Trang (ZRC.MOL.24929; PMBC 25332); Pak Meng, Trang (ZRC.MOL.24067); Phuket, Ao Nam Bor (NHMD 915909; PMBC 3104, 3732); Gulf of Thailand- Pranburi (ZRC.MOL.24930; PMBC 25333); Hua Hin (ZRC.MOL.24004, 24005); Bangsaen (ZRC.MOL.24931, 24932; PMBC 25334); Pattaya (ZRC.MOL.24006); Laem Hin, Trat (ZRC.MOL.24933, 24934; PMBC 25335, 25336); Malaysia: Johor west coast-Sungei Punggur (ZRC.MOL.24935); Sungei Lurus (ZRC.MOL.24936); Pontian Kechil (ZRC.MOL.24937); Minyak Beku (ZRC. MOL.24938); Selangor-Jeram (NHMD 77043); Perak- Teluk Batik, Lumut (ZRC.MOL.24039, 24939); Teluk Senangin (ZRC.MOL.24940); Kedah-Tanjung Jaga (ZRC.MOL.24941); Penang-Gertak Sanggul (ZRC. MOL.24942); Batu Ferringhi (ZRC.MOL.24943); Sarawak-Santubong (ZRC.MOL.24944); Kampung Bako, north of Kuching (ZRC.MOL.24022, 24945); Singapore: Raffles Marina (ZRC.MOL.24946); Tengeh Buoy (ZRC.MOL.24947); Sarimbun (ZRC.MOL.24064); Sungei Buloh (ZRC.MOL.24948, 24949); Kranji (ZRC. MOL.24950); Sembawang Park (ZRC.MOL.24951); Tanjong Irau, Sembawang (ZRC.MOL.24952, 24953); Punggol Point (ZRC.MOL.24053); Changi Village (ZRC.MOL.24056, 24057, 24058); Paku Buoy (ZRC. MOL.24954); Siglap Canal (ZRC.MOL.24955);
Indonesia: Surabaya , East Java ( ZRC.MOL.24956). See also the Supporting Information ( Table S1 View Table 1 ) .
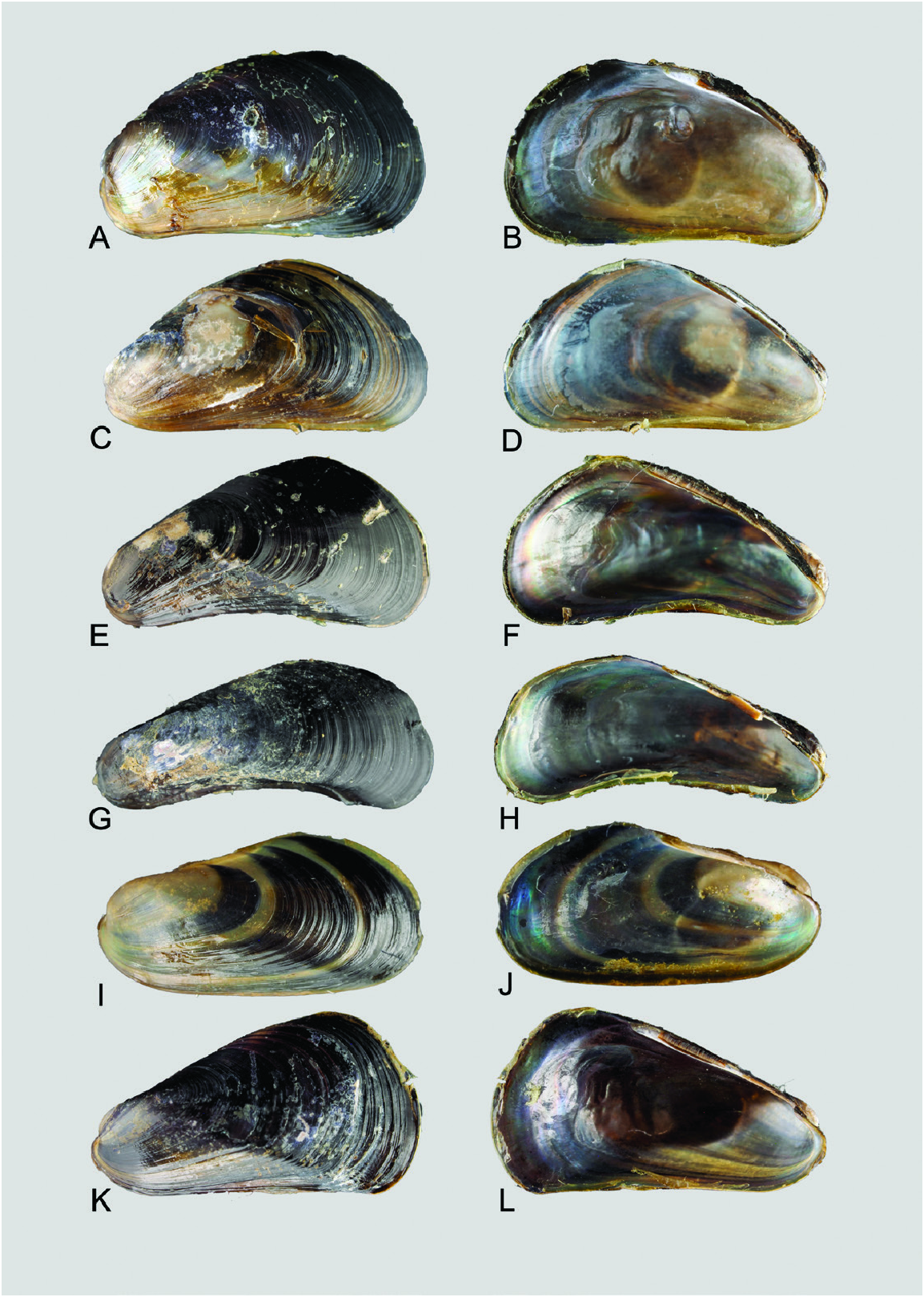
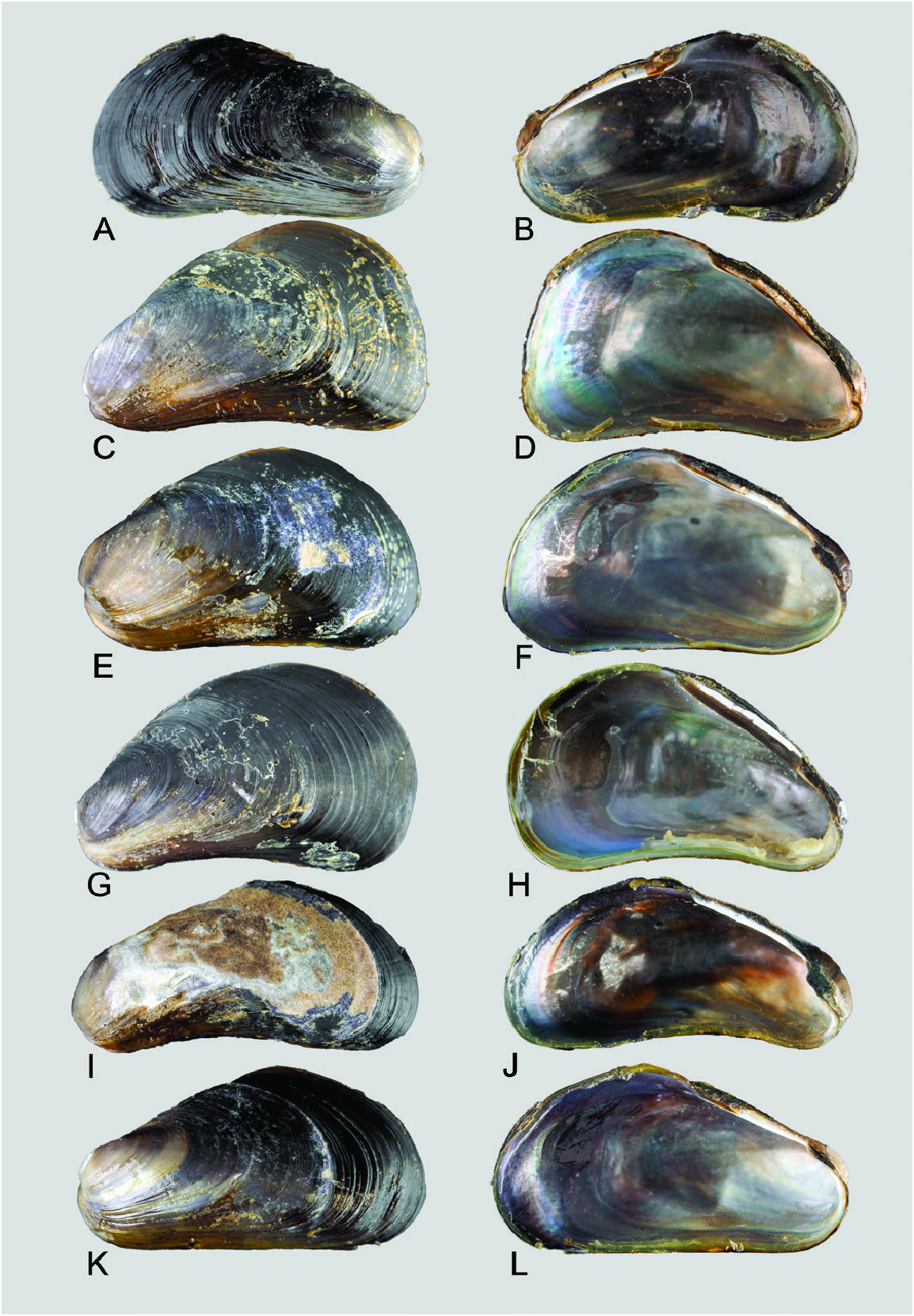
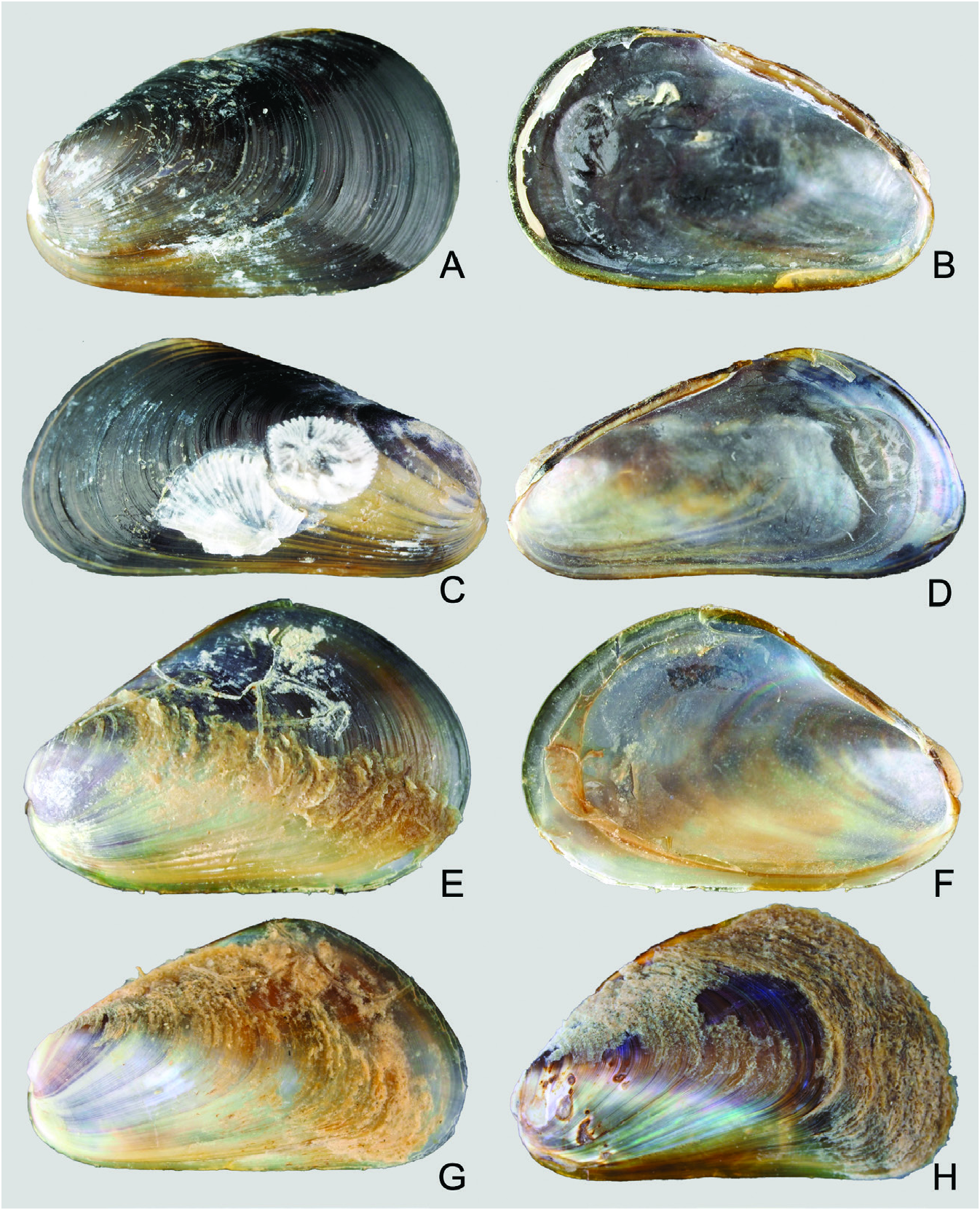
Shell: Variable in outline and shape, length ≤ 15 mm, thin, overall outline trapezoid; the blunt, inflated anterior region is defined by distinct umbonal and anteroventral extensions, while the posterior region is broad and flat. Surface mostly entirely purplish black over the posterior two-thirds of length of valves, while the anteroventral region is translucent yellow to orange crossed by fine dark purple, closely set commarginal lines where the periostracum is intact. Umbones in most cases subterminal to the distinct, rounded anteroventral margin. Shell interior iridescent, margins devoid of teeth. Ligament narrow, smooth without teeth; resilial pits absent.
Shell microstructure: Shell generally comprises two layers below the periostracum: a thin prismatic layer, under which a thick, sheet nacreous layer is present. A thin, fibrous prismatic myostracum may be present within the sheet nacreous layer.
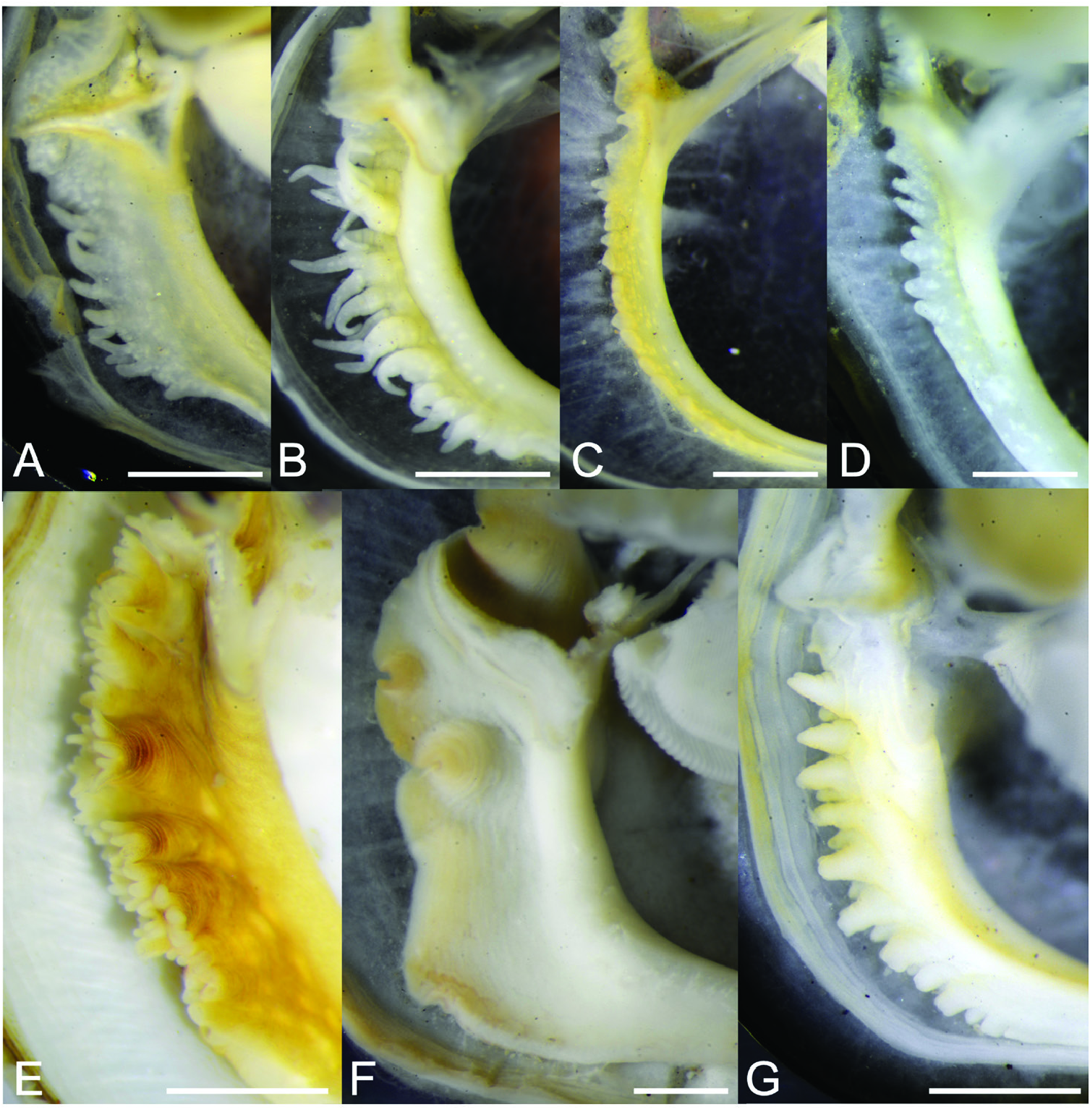
Anatomy: Up to 14 simple or branched (bifid, trifid or quadrifid) guard papillae ( Figs 10E–G View Figure 10 , 12B View Figure 12 ) along one edge of inner mantle forming inhalant siphon. Papillae translucent pink to white, with small white subcutaneous pigment grains. Smaller individuals tend to have fewer guard papillae, which are invariably simple (not branched). In individuals with SLs <2–3 mm, the inner mantle edge forming the inhalant aperture is generally entire, with no papillae. Foot yellowish white, vermiform.
Two adjacent bundles of the posterior adductor muscle can be distinguished. One is generally larger than the other, although their relative size varies between individuals. Posterior byssal retractor muscles are split into two bundles near base of foot, but the anterior bundle is further divided into two or three subbundles towards the shell (as observed on animal, but the muscle scars on the valves do not reflect this separation). Pericardium lies anterior to the posterior byssal retractor muscle complex (Category 2 of Morton, 2015a).
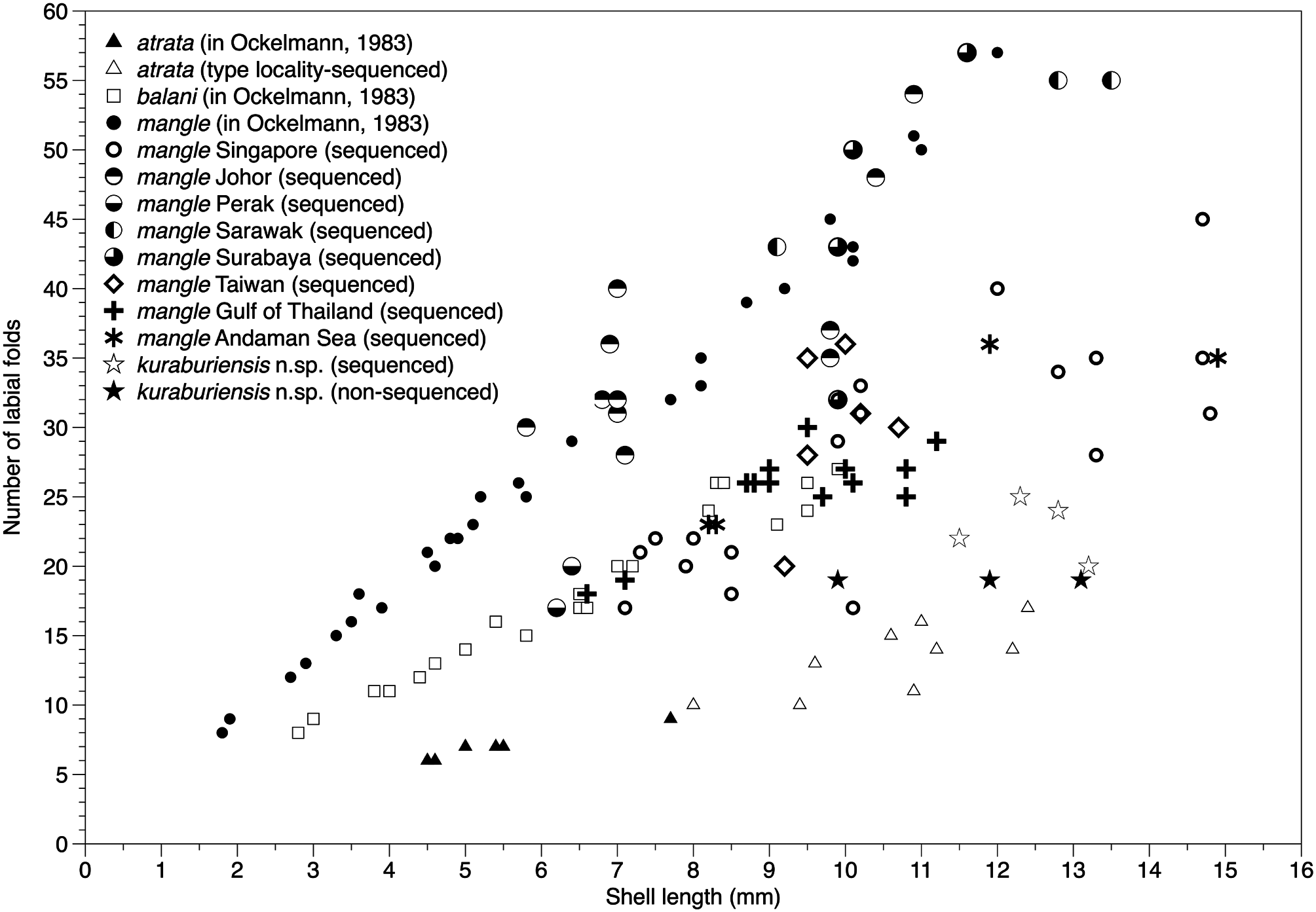
Plicate organ (organ of Sabatier; Fig. 10D View Figure 10 ) present on the roofs of the suprabranchial chamber, along length of bases of outer and inner demibranchs on the left and right sides of the animal. Labial palps vary in length according to shell size (see Theisen, 1982; Ockelmann, 1983), as does the number of folds (sorting ridges), which can range between nine and 55 for SLs 2–13 mm ( Fig. 11 View Figure 11 ).
Ecology: Found singly or in small groups of two to four individuals inside empty barnacle (often Balanus amphitrite Darwin, 1854 ) shells together with Acari, collembolans and the estuarine bryozoan Sundanella sibogae (Harmer, 1915) in the upper- to mid-intertidal zone of estuaries and mangroves. They can also be found in large numbers clustered together over rocks or concrete surfaces in the intertidal zone in estuaries. However, they can also occur subtidally, as observed by Madin et al. (2009) on recruitment nylon net panels placed vertically in floating net cages. Leh et al. (2012) showed that V. mangle , together with other bivalves [ Anadara granosa (Linnaeus, 1758) and Placuna sp. ] composed ≤ 50% of the diet of the eel catfish Plotosus canius Hamilton, 1822 along the west coast of peninsular Malaysia.
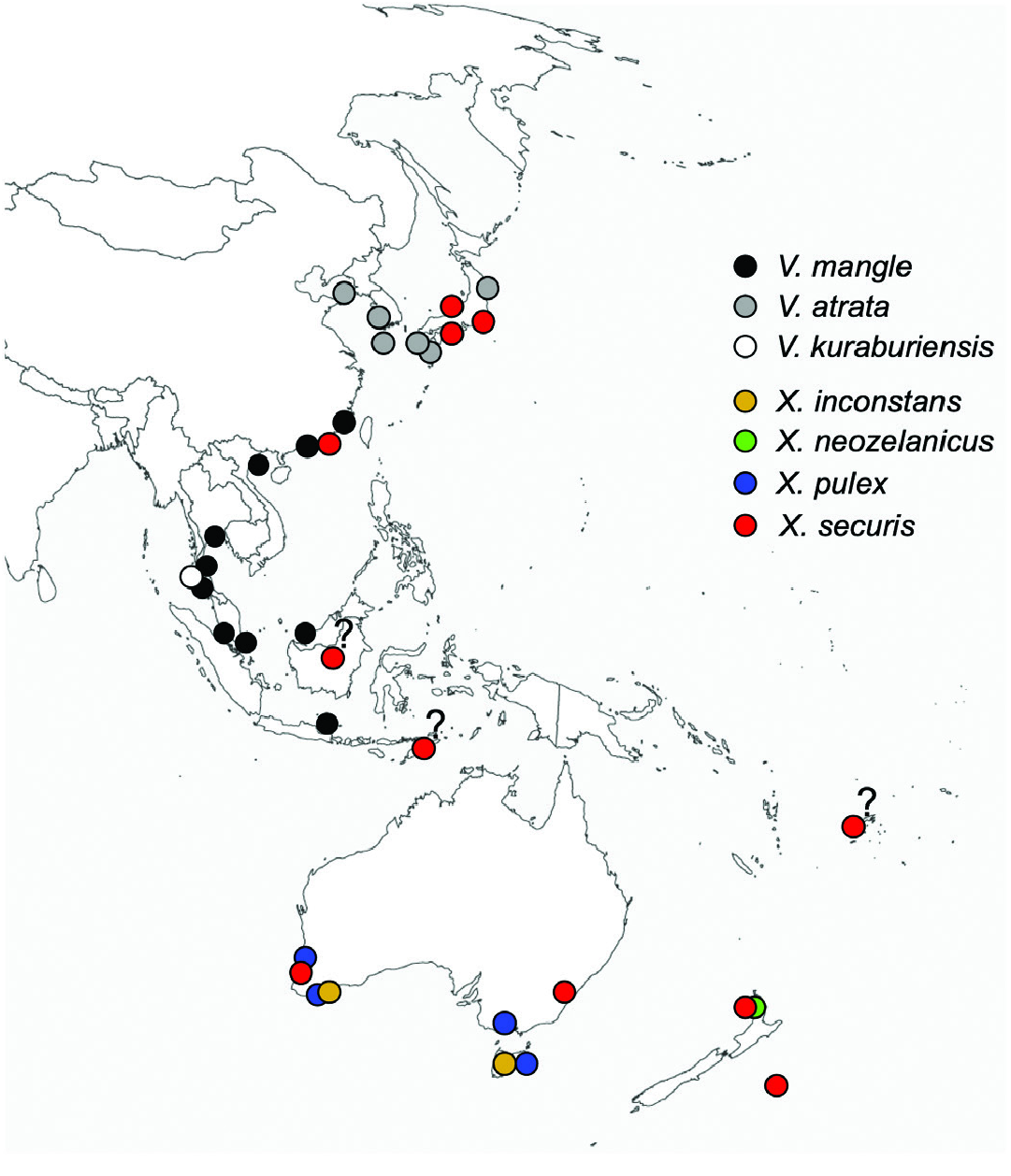
Geographical distribution ( Fig. 13 View Figure 13 ): Kinmen (off Xiamen, China) southwards to the Gulf of Thailand, Malaysia, Singapore, Strait of Malacca and the Andaman Sea; Borneo and Java. Curiously, there were no records of V. mangle or V. atrata from the main island of Taiwan itself in the National Museum of Natural Science in Taichung, Taiwan.
Taxonomic remarks: Ockelmann (1983) described two Xenostrobus species, X. mangle and X. balani , from Jeram in the Malacca Strait and Phuket Island adjoining the Andaman Sea, respectively. They were distinguished chiefly based on the sizes of their shells and labial palps, which are both relatively larger in X. mangle compared with X. balani as diagnosed by Ockelmann ( Table 1 View Table 1 ). Also, size asymmetry of the two bundles making up the posterior adductor muscle was noticed to be more marked in X. balani than in X. mangle . ‘Special byssal secretions’ ( Fig. 10C View Figure 10 ) on the shell surface of X. balani (see paratypes: Fig. 9E, G, H View Figure 9 ) that comprise commarginal thickenings or serrations are weak or absent in X. mangle , according to Ockelmann (1983). In addition, the byssal gland of X. balani is branching and penetrates deeply into the visceral mass, in contrast to the condensed, nonbranching gland in X. mangle (for a summary of characters, see Table 1 View Table 1 ).
In this study, we collected, examined and sequenced material from across a wide geographical range encompassing locations in the East China Sea, South China Sea, Malacca Strait, the Andaman Sea and Java Sea. Our observations suggest the existence of a continuum of morphological character states, instead of disjunct states that Ockelmann used to distinguish X. mangle and X. balani . This is particularly the case for the relative sizes of two muscle bundles of the posterior adductor muscle, the presence or absence of byssal secretions on the valves, the condition of the byssal gland and the number of folds on the labial palps. Nevertheless, the existence of two phenotypes [i.e. individuals having labial palps that are either short or long (corresponding to a smaller or larger number of folds) as shown by Ockelmann (1983)] appears to be real (see Fig. 11 View Figure 11 ) amongst the otherwise genetically indistinguishable individuals we observed in this study. Interestingly, the two phenotypes seem to be location specific; for instance, all individuals collected in this study and sequenced from Singapore, Gulf of Thailand, Andaman Sea and Taiwan had small labial palps, whereas those from localities in Johor, Sarawak (Borneo) and Surabaya in Java had large labial palps. However, they all shared the apomorphy of having a plicate gland along the bases of the outer and inner demibranchs on both left and right sides of the animal.
Ockelmann (1983) also observed that ‘almost all’ X. balani were found singly in empty barnacle shells, whereas X.mangle were‘…living together with Balanus amphitrite …’ and ‘…were found byssally attached to their congeners’ valves’, occurring ‘…gregariously as members of the epifauna on mangrove roots and on stony substrata’. Our observations showed that X. mangle can indeed be found inside barnacle shells, not only singly, but gregariously in groups of up to several individuals depending on the size of the empty barnacle shells. These tended to be smaller individuals of X. mangle possessing largely intact ‘special byssal secretions’ ( Fig. 10C View Figure 10 ), possibly because of the sheltered nature of the microhabitat, which is also used by other organisms, such as mites, collembolans and the high shore bryozoan Sundanella sibogae . Conversely, Ockelmann (1983) stated that many individuals of X. mangle living outside barnacle shells had live barnacles attached to their shells, which he attributed to the absence of ‘special byssal secretions’. We also observed some individuals fouled by barnacles, but these were not restricted to those living outside dead barnacle shells. Hence, based on these field observations, considered together with their similar morphology and genetic composition, we conclude that X. balani and X. mangle are one and the same species.


| ZRC |
Zoological Reference Collection, National University of Singapore |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
SuperFamily |
Mytiloidea |
|
Family |
|
|
SubFamily |
Xenostrobinae |
|
Genus |
Vignadula mangle
| Tan, Koh Siang, Tan, Samuel Hui Ming, Sanpanich, Kitithorn, Duangdee, Teerapong & Ambarwati, Reni 2022 |
Xenostrobus aratus (Dunker, 1856)
| Nguyen C 2001: 417 |
Xenostrobus sp.
| Ng PKL & Sivasothi N 1999: 119 |
Xenostrobus atratus
| Kurozumi T 2017: 1173 |
| Kurozumi T 2000: 429 |
| Bernard FR & Cai Y-Y & Morton B 1993: 33 |
Xenostrobus atrata (Lischke, 1871)
| Lee SY & Morton B 1985: 54 |
Xenostrobus mangle
| Ockelmann KW 1983: 106 |
Xenostrobus balani
| Ockelmann KW 1983: 107 |