
Homolophus almasyi ( Roewer, 1911 ) Snegovaya & Cokendolpher, 2021
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4908.3.1 |
|
publication LSID |
lsid:zoobank.org:pub:F2774810-3C77-426E-A1BC-0F42F21E7F9B |
|
DOI |
https://doi.org/10.5281/zenodo.4450788 |
|
persistent identifier |
https://treatment.plazi.org/id/039C113F-0972-FF8F-98EB-DB59FDC1FD58 |
|
treatment provided by |
Plazi |
|
scientific name |
Homolophus almasyi ( Roewer, 1911 ) |
| status |
comb. nov. |
Homolophus almasyi ( Roewer, 1911) , new combination
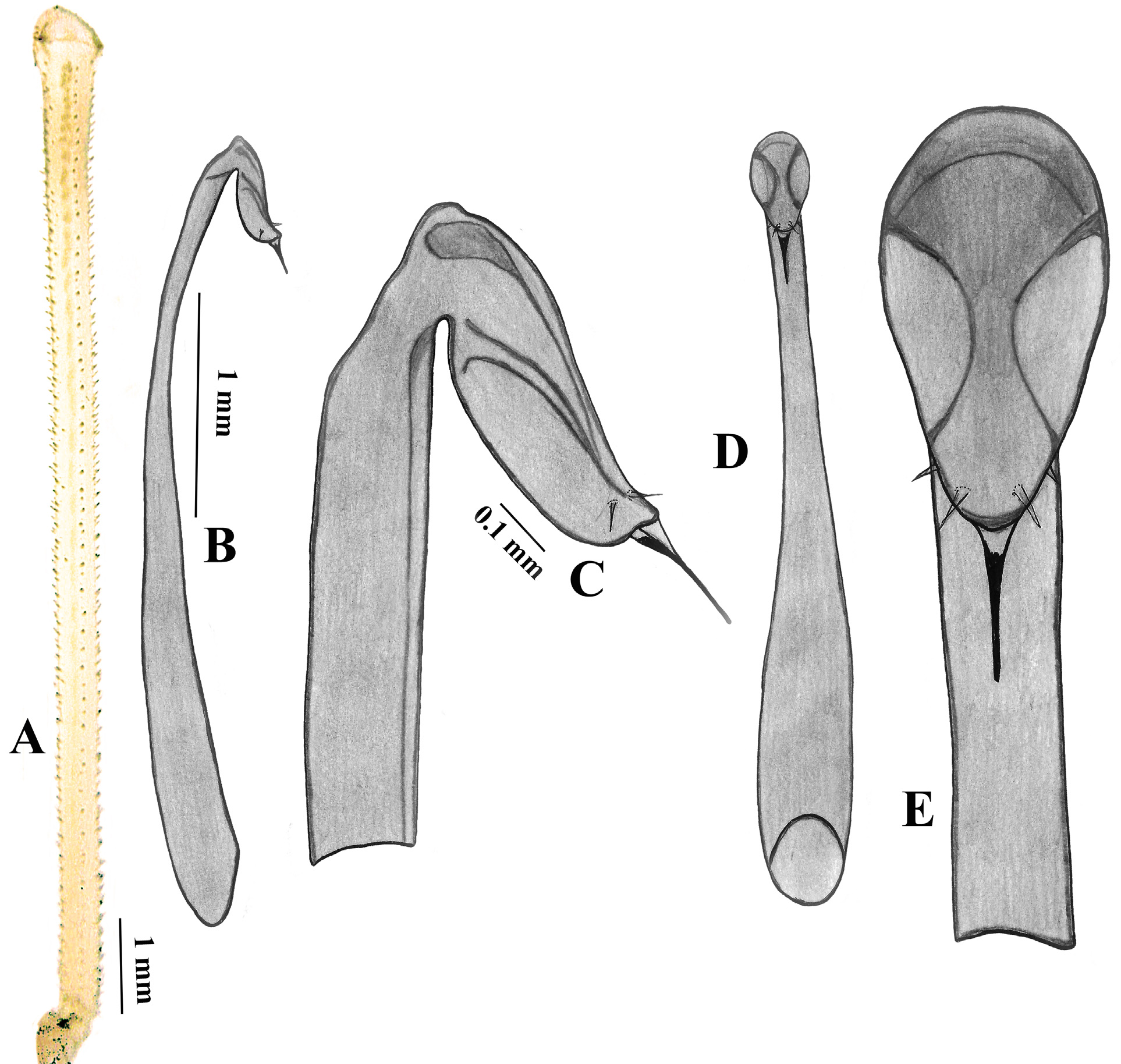
Figures 5–7 View FIGURE 5 View FIGURE 6 View FIGURE 7
Opilio Almasyi Roewer, 1911: 44–45 View in CoL .
Opilio almasyi View in CoL — Šilhavý 1967: 475–478, figs. 14–20; Gricenko 1979a: 35, fig. 28; Gricenko 1980: 556–557
Diagnosis. This species is considered by some as just a color morph of H. asiaticus ( Gricenko, 1979a) , see under comments for the current species. The only difference we can see illustrated in literature is that H. almasyi has a smooth cheliceral basal segment and that segment on H. asiaticus is ventrally with a blunt rounded bulge. Homolophus almasyi differs from all other species including the most closely related, H. silhavyi Staręga & Snegovaya, 2008 , by having the following characters: very long and thin legs (Fe II more than 2.5 times longer than body length); both palpal femur and tibia ventrally with larger and more numerous denticles and granules; penis slender, truncus of penis slowly becoming narrower to apex, glans with recess ventrally on each side.
Type locality. Originally reported ( Roewer, 1911) in German as “Turkestan, Tekkes und Ala-Buga”. These two localities are now known ( Tsurusaki, Tchemeris & Logunov, 2000) as Kazakhstan, Almaty Area, Raiymbek District, Tekes (ca. 42°50’N, 80°03’E) and Kyrghyzstan, Naryn Area, Ak-Talinsky District, Alabuga River (ca. 41°14’N, 74°30’E).
Distribution. Russia, Kazakhstan ( Fig. 7 View FIGURE 7 ).
Specimens Examined. RUSSIA, Gorno-Altaysk (formerly Oirot-Tura ), in gardens of city, on the fence and plants, 30.06.1939, leg. D. Kharitonov (1 male, RCNS) . KAZAKHSTAN, coast of Black Irtysh, against YuzAgach (natural boundary Zhuzagash ) [8 km below Boran (old name Buran)], in a dry potato field, Zaisan expedition of PSU [Perm State University], 17.07.1936, leg. D. Kharitonov, field # Х-28 (1 male, 2 females, RCNS) ; same locality, 27.07.1936, leg. D. Kharitonov, field # Х-83 (1 male, 2 juv., RCNS) ; same locality, the left bank of Black Irtysh, against Yuz-Agach (near Boran ), by the fire at night, 17.07.1936, leg. D. Kharitonov, field # Х-29 (1 male, RCNS) ; same locality, shore of Black Irtysh , by fire, 17.07.1936, leg. D. Kharitonov, field Ñ Х-90 (12 females, PSU / ZMMU) ; same locality, in Altai hut, in room, 6.08.1936, leg. D. Kharitonov, field # Ñ Х-129 (1 male, PSU / ZMMU) ; same locality, Topolev cape, in field, 11.08.1936, leg. D. Kharitonov, field Ñ Х-135 (1 male, PSU / ZMMU) ; same locality, Boran , at school, 27.07.1936, leg. D. Kharitonov, field Ñ Х-84 (1 female, PSU / ZMMU) ; Merkensky district , Almaty (previously Alma-Ata) environs, vic. Podgornoye vill. (now Kirgizsay ), 20.07.1929, field Ñ 61-1937 (1 male, 2 females in 1 vial; 1 male, 2 females in 2 nd vial; CNS) ; Almaty, in clods, 15.07.1935, field Ñ61-1937 (1 female, RCNS) ; Almaty environs, Glubokaya schel” ( Deep gap), 26.06.1936, leg. V. Shandurov (10 juv., RCNS) ; Almaty environs, 3.07.1936, field Ñ61-1937 (1 female, 1 juv., IZB) ; Almaty environs, Talgarsky district, near Medeo , 3– 28.07.1997, leg. A. V. Gromov (2 males, 1 female, RCNS) . Almaty region, Taldy-Kurgan district, Gavrilovka village , 19.09.1930, leg. Ivanov (3 males, RCNS) ; Malaya Almaatinka river , leg. Ivanov, 19.09.1930, field Ñ61-1937 (3 males, RCNS) .
Description. Large harvestmen; male body rectangular, with rounded corners; not granulated, medium sclerotization, slender, 1.87 times longer than wide; length 8.6 mm, width 4.6 mm. Body light brown with darker and lighter spots all over; light color to white, longitudinal stripe (thin, less than ocularium width) extending entire length of body. Along borders of tergites, there are rows of small denticles. Ocularium low, slightly wider than long, not large, 1.2 times its length from anterior margin of cephalothorax, with small 13–14 black-tipped denticles on each side; pair of slightly larger white, with dark-tipped, denticles on anterior border. In front of ocularium and sides of cephalothorax with only few dark tipped denticles. Legs very long, femora to tibia cylindrical in cross-section, slender not thickened, with many denticles at edges, longitudinal rows of denticles on each segment; lengths (mm): I 11.5 + 2.4 + 8.5 + 12.0 + 14.8 = 49.5, II 23.1 + 3.0 + 20.5 + 16.5 + 22.5 = 85.6, III 10.5 + 1.8 + 7.6 + 13.0 + 15.6 = 48.5, IV 13.6 + 2.2 + 9.6 + 16.4 + 22.0 = 63.8. Pedipalp segment lengths (mm): 2.0 + 1.0 + 1.2 + 2.5 = 6.7. Pedipalp not thickened, denticles on Fe dorsally and ventrally, Pa dorsally and laterally, Ti all, Ta ventrally with strip (4–5 denticles wide) of sensory microdenticles. Chelicerae slender with few light-colored tabby marks on distal segment; basal segment dorsally with few smaller denticles, distal segment dorsally with few microdenticles. Cheliceral basal segment ventrally smooth without bulge, length 2.3 mm, distal length 2.8 mm. Penis long, thin; truncus slightly wider at base, dorsoventrally flattened, bowed, recurved in lateral view, lengths (mm): truncus 3.85, glans 0.5, stylus 0.15. Glans with recess ventrally on each side.
Female differs from male by having wider abdomen and absence of armaments on chelicerae and pedipalps; less slender, 1.94 times longer than wide; length 7.0 mm, width 3.6 mm. Leg lengths (mm): I 7.0 + 1.8 + 5.2 + 6.5 + 10.5 = 31.0, II 13.4 + 2.0 + 10.6 + 9.0 + 23.6 = 58.6, III 6.0 + 1.6 + 5.0 + 7.4 + 11.0 = 31.0, IV 9.0 + 2.0 + 7.0 + 11.0 + 16.0 = 45. Pedipalp segment lengths (mm): 1.3 + 0.7 + 1.0 + 1.9 = 4.9. Cheliceral basal segment length 1.6 mm, distal segment length 1.8 mm.
Comments. Specimens over 80 years old. Colors likely faded. In the past [ Staręga (1978, 2003), Gricenko (1979a, 1980), Tchemeris et al. 1998, and Tsurusaki et al. (2000 a)] there has been confusion on the identification and generic placement of this species ( almasyi Roewer, 1911 ), asiaticus Gricenko, 1979a , and potanini Simon, 1895 . Staręga (1978) and Tsurusaki et al. (2000), considered Opilio almasyi and Opilio asiaticus both as junior subjective synonyms of Phalangium potanini . This situation remained until Snegovaya (2018) removed potanini as the type species of the new genus Gricenkovia but made no comment on the two other species. Chemeris et al. 1998 noted that “It is very likely that Opilio asiaticus is only a colour morph of Opilio almasyi Roewer, 1911 [for distinguishing colour characters see Gritsenko, 1979.]” They further remarked “The problem is open until more specimens, including the types of both species, are studied.” We completely agree and having not studied any types, retain the two as separate species for now.


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Opilioninae |
|
Tribe |
Opilionini |
|
Genus |
Homolophus almasyi ( Roewer, 1911 )
| Snegovaya, Nataly Yu. & Cokendolpher, James C. 2021 |
Opilio almasyi
| Gricenko, N. I. & H. I 1980: 556 |
| Gricenko, N. I. & H. I 1979: 35 |
| Silhavy, V. 1967: 475 |
Opilio Almasyi Roewer, 1911: 44–45
| Roewer, C. F. 1911: 45 |