
Colletes jankowskyi Radoszkowski, 1891
|
publication ID |
https://doi.org/ 10.17161/jom.v0i63.5672 |
|
DOI |
https://doi.org/10.5281/zenodo.8132282 |
|
persistent identifier |
https://treatment.plazi.org/id/039C0042-FF98-5525-FE68-D8F6797CA3E0 |
|
treatment provided by |
Felipe |
|
scientific name |
Colletes jankowskyi Radoszkowski, 1891 |
| status |
|
Colletes jankowskyi Radoszkowski, 1891 View in CoL View at ENA
( Figs. 1, 2 View Figures 1–2 , 4 View Figures 3–4 , 7 View Figures 5–7 , 9 View Figures 9–13 )
Colletes jankowskyi Radoszkowski, 1891: 253 View in CoL .
Colletes ventralis Pérez, 1903: 225 . Synonymy vide Kuhlmann, 2000: 180.
Colletes ventraliformis Cockerell, 1924: 595 . Synonymy vide Kuhlmann, 2000: 180.
SPECIMENS EXAMINED: [ JAPAN] Honshu: 1♂, Karuizawa ( Kitasaku-dist. ), Nagano Pref., 30.viii.1949 [30 August 1949] ( R. Ishikawa, ELKU). Kyushu: 1♂, Kujû ( Bungo ), 3.viii.1940 [3 August 1940] ( T. Esaki, ELKU; a paratype of C. babai ) ; 1♀, 2♂♂, Kokonoe-machi, Kusu-gun, Kuju , Oita Pref., 13.ix.2009 [13 September 2009] (O. Tadauchi, ELKU) ; 1♀, 1♂, Chojyabaru, alt. 1,050 m, Kokonoe-machi, Kusu-gun , Kuju , Oita Pref., N33°7’6.773’’ E131°13’49.331’’, 5.ix.2010 [5 September 2010] ( R. Murao, on Lespedeza cyrtobotrya Miq. , cMur) GoogleMaps ; 2♀♀, 7♂♂, same locality, 28.viii.2011 [28 August 2011] ( R. Murao & Y. Murao, on L. cyrtobotrya, ELKU ) GoogleMaps ; 6♀♀, same locality, 31.viii.2012 [31 August 2012] ( R. Murao, on L. cyrtobotrya, ELKU ) GoogleMaps ; 4♀♀, same locality, 17.ix.2014 [17 September 2014] ( R. Murao, on L. cyrtobotrya, ELKU ) GoogleMaps ; 2♀♀, 4♂♂, same locality, 15.viii.2015 [15 August 2015] ( R. Murao, on L. cyrtobotrya, ELKU ) GoogleMaps ; 14♀♀, 2♂♂, same locality, 28.viii.2015 [28 August 2015] ( R. Murao, on L. cyrtobotrya, ELKU ) GoogleMaps ; 19♀♀, same locality, 5.ix.2015 [5 September 2015] ( R. Murao, on L. cyrtobotrya, ELKU ) GoogleMaps ; 2♀♀, same locality, 14.ix.2015 [14 September 2015] ( R. Murao, on L. cyrtobotrya, ELKU ) GoogleMaps .
DIAGNOSIS: In Japan, this species is similar to C. babai and C. floralis . It can be differentiated from these species by the following key:
1. ♀♀ ................................................................................................................................. 2
—. ♂♂ ................................................................................................................................. 4
2(1). Malar space long, approximately one-half of basal width of mandible; punctures on 1 st metasomal tergum sparser, as in figure 3 (IS = 3d at maximum) ... ................................................................................................... C. floralis Eversmann View in CoL
—. Malar space short, less than half of basal width of mandible; punctures on 1 st metasomal tergum denser, as in figure 4 (IS = 0.5–2d) .......................................... 3
3(2). Mesoscutum with yellowish-brown hairs, excluding darker setae ...................... .................................................................................. C. babai Hirashima & Tadauchi View in CoL
—. Mesoscutum with whitish hairs excluding dark setae ........................................... ....................................................................................... C. jankowskyi Radoszkowski View in CoL
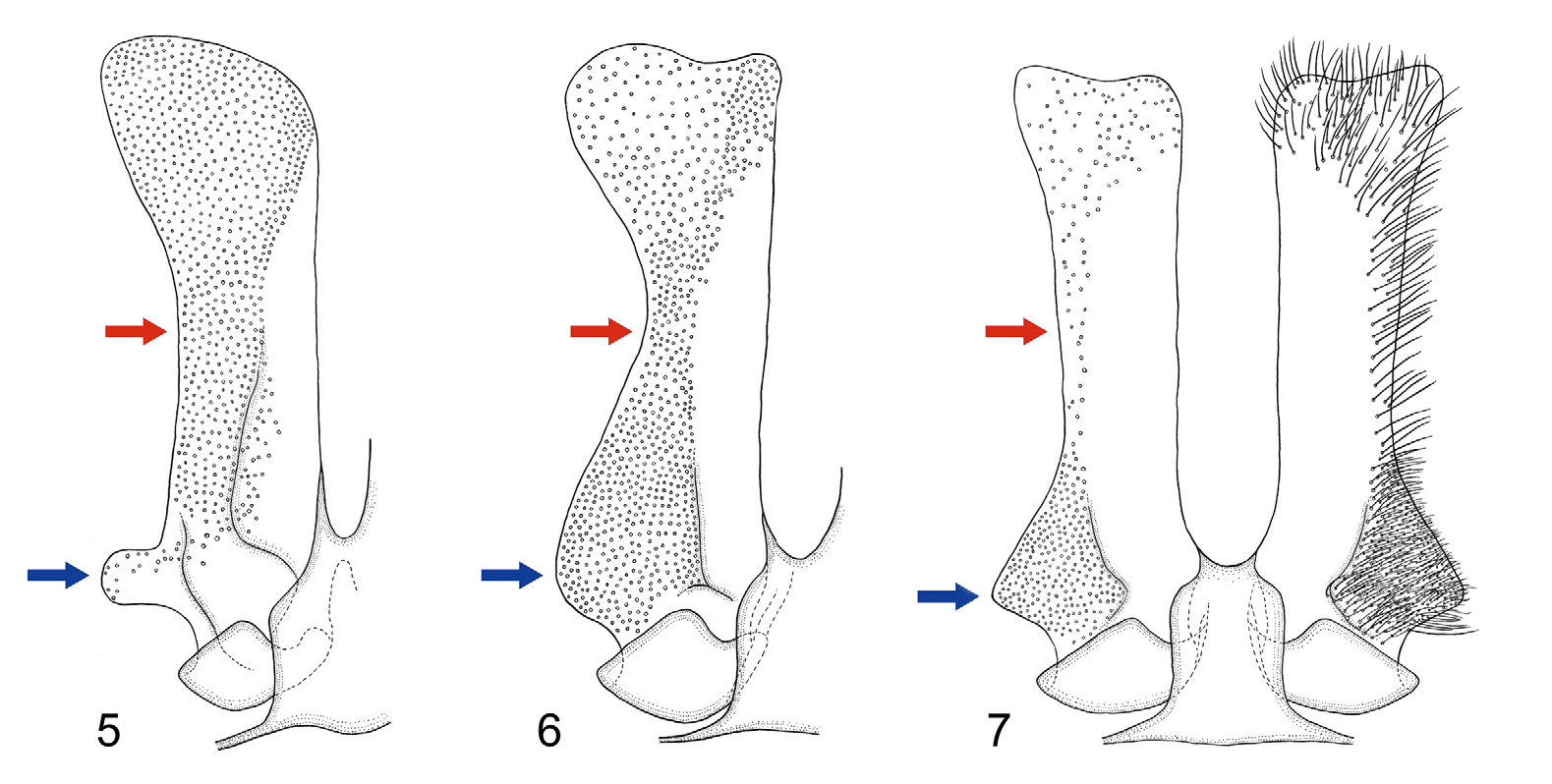
4(1). Pilosity of mesoscutum and mesoscutellum without dark hairs; base of 7 th metasomal sternum with lateral process (blue arrow in Fig. 5 View Figures 5–7 ); hair zone on disc of 7 th metasomal sternum occupies over half of width of disc in central area (red arrow in Fig. 5 View Figures 5–7 ) .............................................................. C. floralis Eversmann View in CoL
—. Pilosity of mesoscutum and mesoscutellum mixed with dark hairs; base of 7 th metasomal sternum laterally rounded or angulate (blue arrow in Figs. 6–7 View Figures 5–7 ); hair zone on disc of 7 th metasomal sternum occupies approximately half or less than half of width of disc in central area (red arrow in Figs. 6–7 View Figures 5–7 ) .... 5
5(4). Base of 7 th metasomal sternum laterally rounded (blue arrow in Fig. 6 View Figures 5–7 ); hair zone on disc of 7 th metasomal sternum occupies approximately half of width of disc in central area (red arrow in Fig. 6 View Figures 5–7 ) ......... C. babai Hirashima & Tadauchi View in CoL
—. Base of 7 th metasomal sternum laterally angulate (blue arrow in Fig. 7 View Figures 5–7 ); hair zone on disc of 7 th metasomal sternum narrow, occupies only outer margin of disc in central area (red arrow in Fig. 7 View Figures 5–7 ) ........... C. jankowskyi Radoszkowski View in CoL
DISTRIBUTION: Japan (central Honshu and Kyushu); Russia, Mongolia, China, Tajikistan ( Proshchalykin & Kuhlmann, 2015).
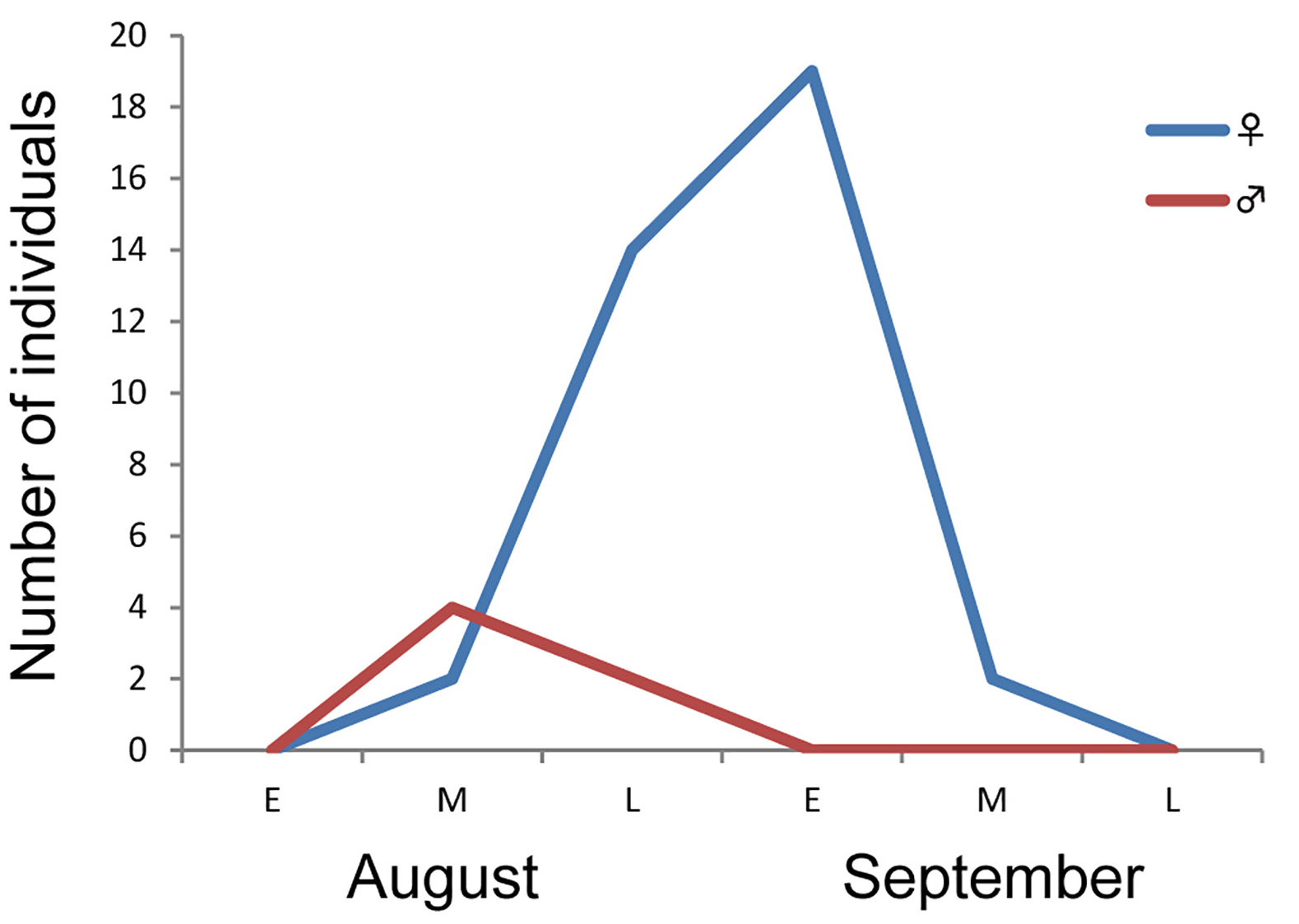
PHENOLOGY IN JAPAN: The phenology based on the data of both the field survey and ELKU collection is as follows: females emerged from mid-August to mid-September and males from early August to mid-September. The emergence of both females and males could not be confirmed in other seasons. Thus, C. jankowskyi seems to be a univoltine species in Japan. Figure 8 View Figure 8 shows the phenology of C. jankowskyi by number of individuals sampled in Oita Prefecture, Kyushu, in 2015. The number of female individuals was highest from late August to early September and distinctly decreased thereafter.
HABITAT IN JAPAN: This species is found in semi-natural grassland ( Fig. 10 View Figures 9–13 ) at an elevation of approximately 1000 m.
FLORAL RECORDS IN JAPAN: The flower records obtained from the field survey were from three species in three families: Aralia cordata Thunb. (Araliaceae) ( Fig. 12 View Figures 9–13 ), Echinops setifer Iljin (Asteraceae) ( Fig. 13 View Figures 9–13 ), and L. cyrtobotrya (Fabaceae) ( Fig. 11 View Figures 9–13 ). The flowers most frequently visited by C. jankowskyi were those of L. cyrtobotrya (96♀♀, 15♂♂: 50♀♀, 14♂♂ by sampling, and 46♀♀, 1♂ from observations at flowers), followed by A. cordata (5♀♀, 1♂ from observations at flowers), and E. setifer (1♂ from observation at flowers). During the periodical sampling conducted from August to September 2015, C. jankowskyi visited only L. cyrtobotrya (37♀♀, 6♂♂), despite the presence of many other flowers (44 species) in bloom during the active season of C. jankowskyi . At least at the study site, the population of C. jankowskyi appears to have a floral preference for L. cyrtobotrya .
DNA BARCODES: The pair-wise sequence divergence was 7.5–7.8% between C. jankowskyi and C. floralis and 2.9–3.5% between C. jankowskyi and C. babai . The intraspecific divergence was 0.3–0.7% (inter- and intraspecific divergence summarized in Table 2 View Table 2 ). Thus, DNA barcoding data can be used for discrimination of C. jankowskyi from both C. babai and C. floralis .



| R |
Departamento de Geologia, Universidad de Chile |
| T |
Tavera, Department of Geology and Geophysics |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Colletes jankowskyi Radoszkowski, 1891
| Murao, Ryuki, Ikudome, Shuichi & Tadauchi, Osamu 2016 |
Colletes ventraliformis
| Kuhlmann, M. 2000: 180 |
| Cockerell, T. D. A. 1924: 595 |
Colletes ventralis Pérez, 1903: 225
| Kuhlmann, M. 2000: 180 |
| Perez, J. 1903: 225 |
Colletes jankowskyi
| Radoszkowski, O. 1891: 253 |