
Seira picoensis, Nunes & Cipola & Bellini, 2021
|
publication ID |
https://doi.org/10.11646/zootaxa.5048.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:D8D39DE7-B270-4D1D-8CB3-25286966F751 |
|
persistent identifier |
https://treatment.plazi.org/id/039B87F0-D704-FC71-FF49-F89EBD96FD7B |
|
treatment provided by |
Plazi |
|
scientific name |
Seira picoensis |
| status |
sp. nov. |
Seira picoensis View in CoL sp. nov. Nunes, Bellini & Cipola
Figs 2–12 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 View FIGURE 7 View FIGURE 8 View FIGURE 9 View FIGURE 10 View FIGURE 11 View FIGURE 12 , Table 1
Type material. Holotype female on slide ( CC /UFRN), Brazil, Piauí State, Picos municipality, vicinities of Instituto Federal do Piauí ( 7°3’57.71”S, 41°24’2.13”W), Caatinga biome, 13.ii.2017, R. C. Nunes coll. 4 paratypes on slides: GoogleMaps
1 female, same data as holotype ( INPA-CLL 0000115 ) GoogleMaps ; 2 females and 1 male ( CC /UFRN), same data as holotype, except for Picos municipality, road BR-407 towards Jaicós municipality ( 7°15’14.06”S, 41°15’21.60”W) GoogleMaps .
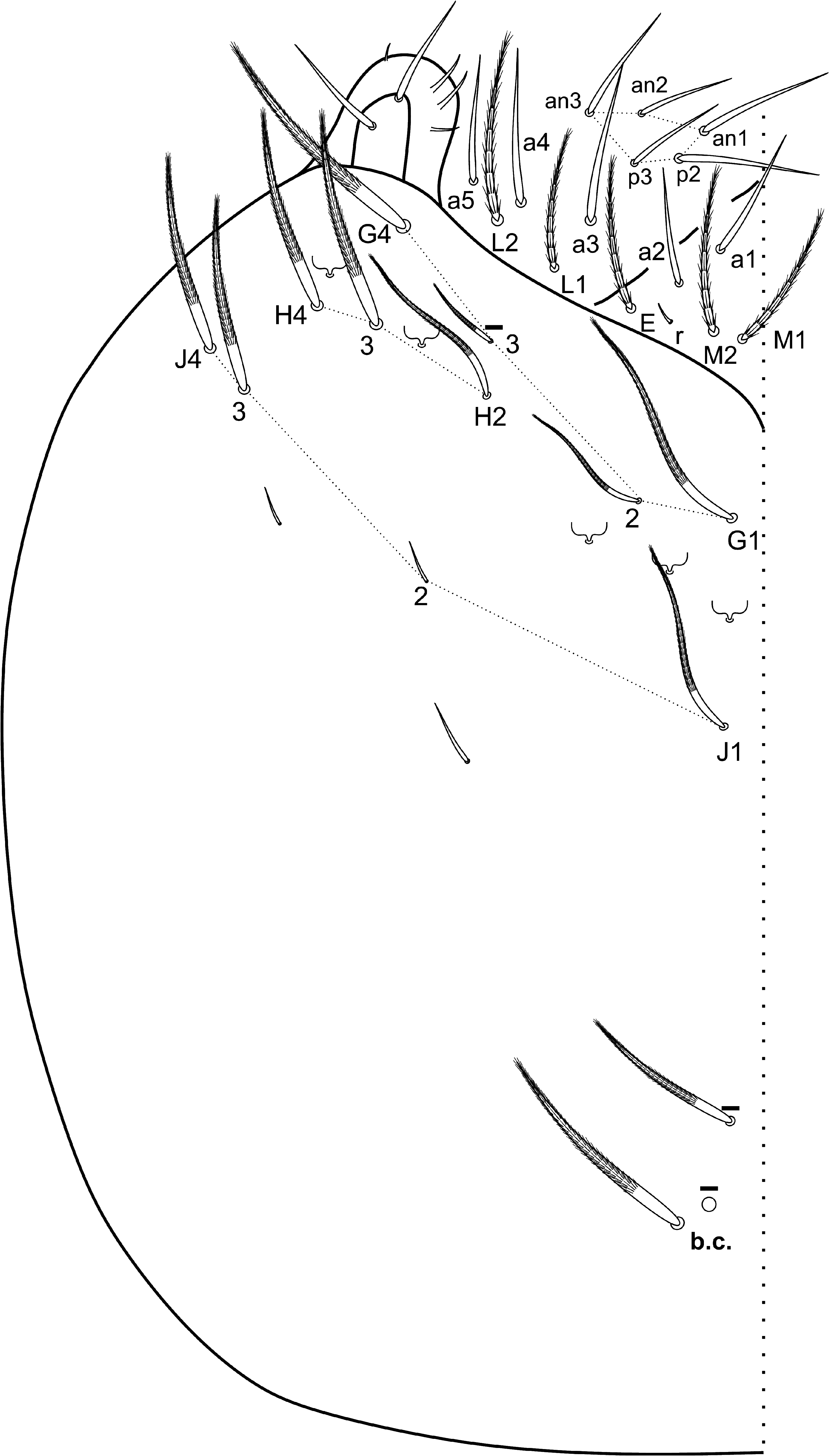
Diagnosis. Ant. IV not annulated or subsegmented, with a bilobed apical bulb. Two conic inner labral papillae, outer papillae smaller and almost imperceptible, as small protuberances. Labial basomedian field with r chaeta reduced. Head’s ventral groove surrounded by 3–5 ciliated chaetae, post-labial chaetotaxy with G1–4 ( G3 may be absent), H2–4, J1–4, all ciliated with exception of J2, plus 2 small smooth chaetae. Dorsal head macrochaetotaxy with 7–8 ‘An’, 4–5 ‘A’ ( A0, A2–5; A4 mac or mic), 3 ‘M’ ( M1–2, M4), 5 ‘S’ ( S0–3, S5), 5 ‘Pa’ ( Pa1–5), 2 ‘Pm’ ( Pm1, Pm3), 4 ‘Pp’ ( Pp1–3, Pp5), 1–2 ‘Pe’ ( Pe2–3, Pe2 present or absent) plus one posterior mac without clear homology. Th. III with 11 ( a1a, a1–5, p1–2–2a, p2ea and p3) and Abd. I with 4 ( m2–2i, m3 and m4i) central mac, respectively. Abd. IV with 7–8 central mac ( A3p, A5, B3–6, Si and Sm), with 3–6 posterior psp. Metatrochanteral organ with 12–22 spiny smooth chaetae. Unguiculus acuminate, with the postero-external lamella serrated. Collophore anterior side with 3 distal ciliated mac plus at least 3 proximal spines, posterior side with 1 smooth chaeta plus 3–4 short spines, lateral flap with about 10 smooth and 4 ciliated chaetae. Manubial plate with 4 ciliated chaetae and 2–3 psp. Manubrium ventral formula with 1,2,2,2/2,12 ciliated chaetae.
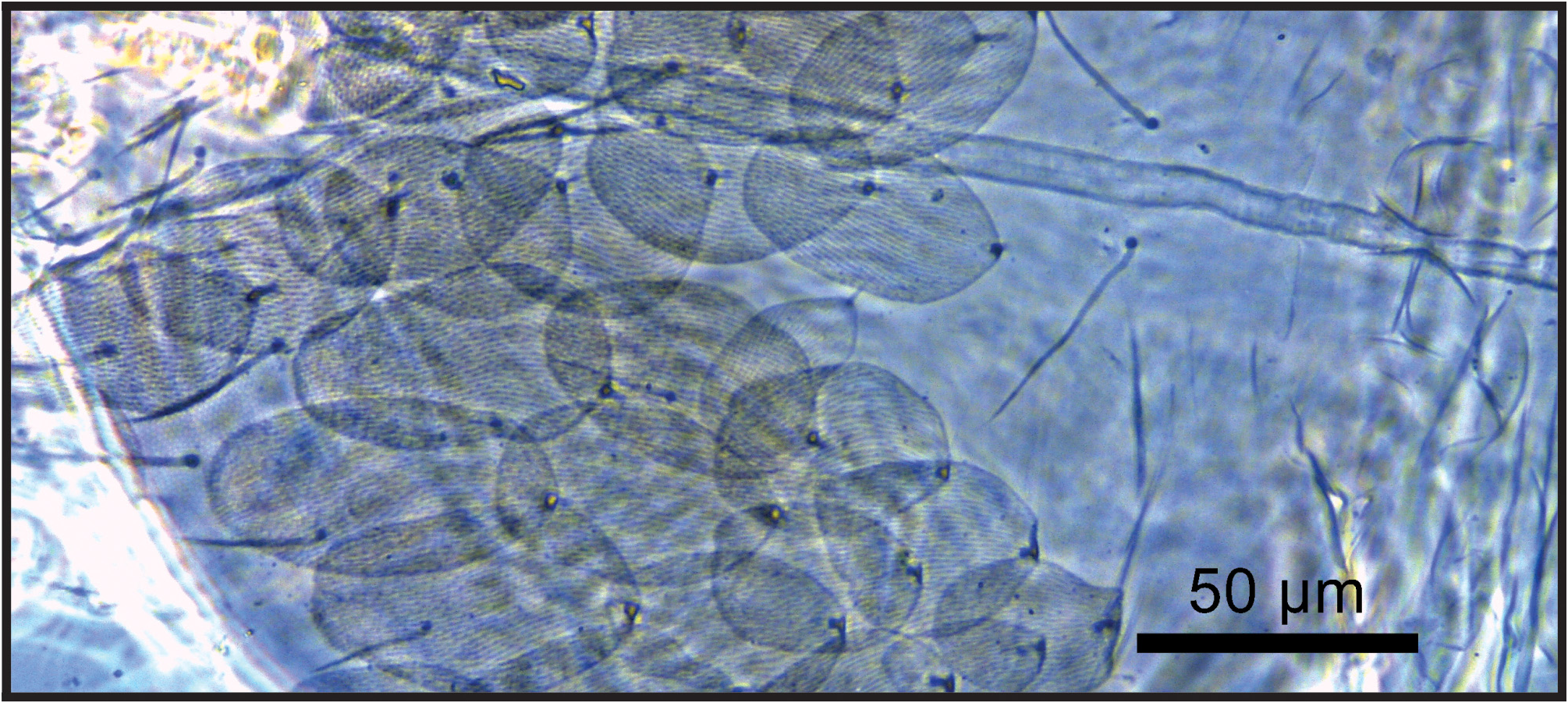
Description. Habitus as in Fig. 2 View FIGURE 2 . Pale to yellowish ground with dark and light blue pigments on Ant. I–IV, anterior dorsal head (between the eyes), light blue pigment over legs and anterolateral edges of the Th. II. Eye patches deep black. Scales covering Ant. I–II, proximal 1/3 of Ant. III, all head, trunk, legs (except empodia), anterior collophore (proximally), manubrium and dens ventrally. Total length (head + trunk) of type series ranging between 1.15–1.82 mm in adults ( holotype 1.68 mm). Antennae shorter than body length. Antennae: trunk ratio as 1: 2–2.57 ( holotype 1: 2.28). Antennal ratio in type series as I: II: III: IV = 1: 1.52–1.68: 1.54–1.97: 2.1–2.81 ( holotype 1: 1.52: 1.54: 2.72). Abd. III: IV ratio in the midline of type series as 1: 3.01–4.38 ( holotype 1: 3.39).
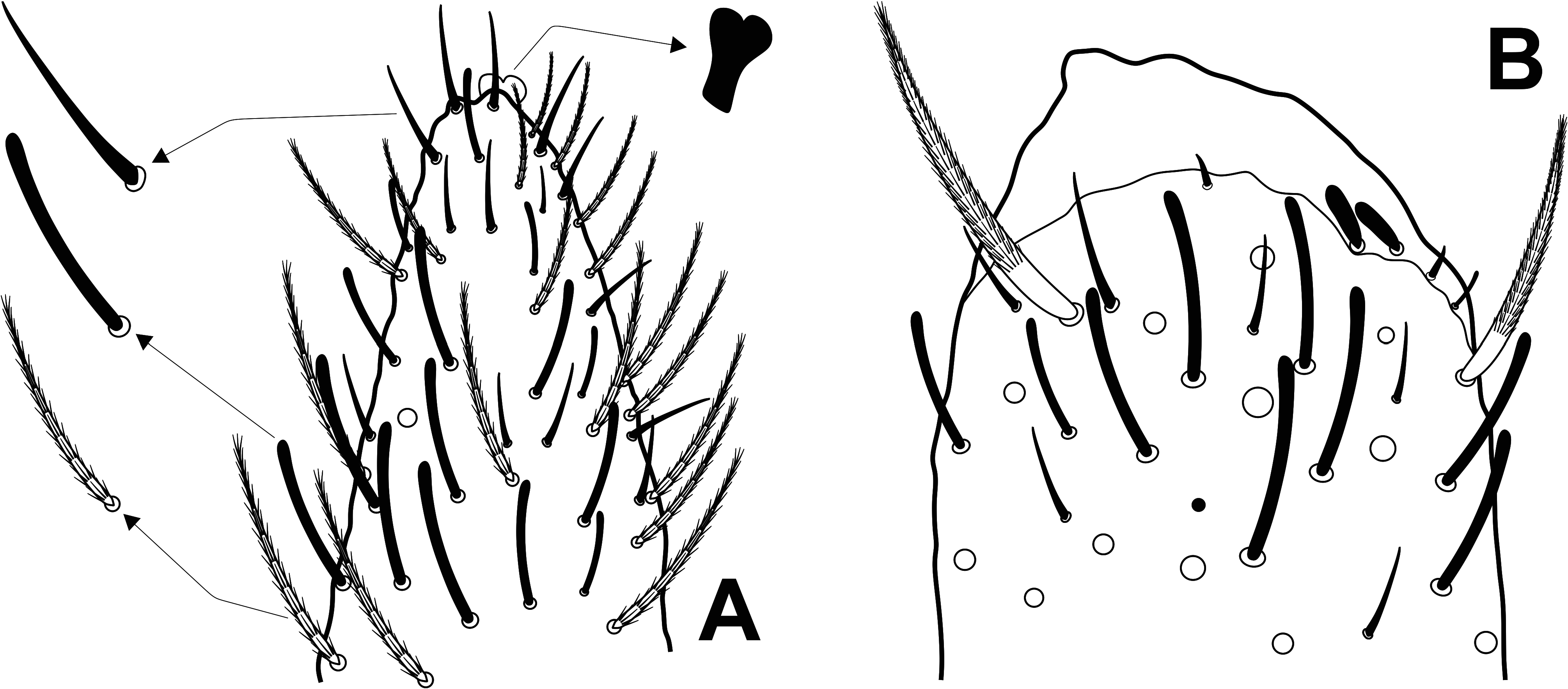
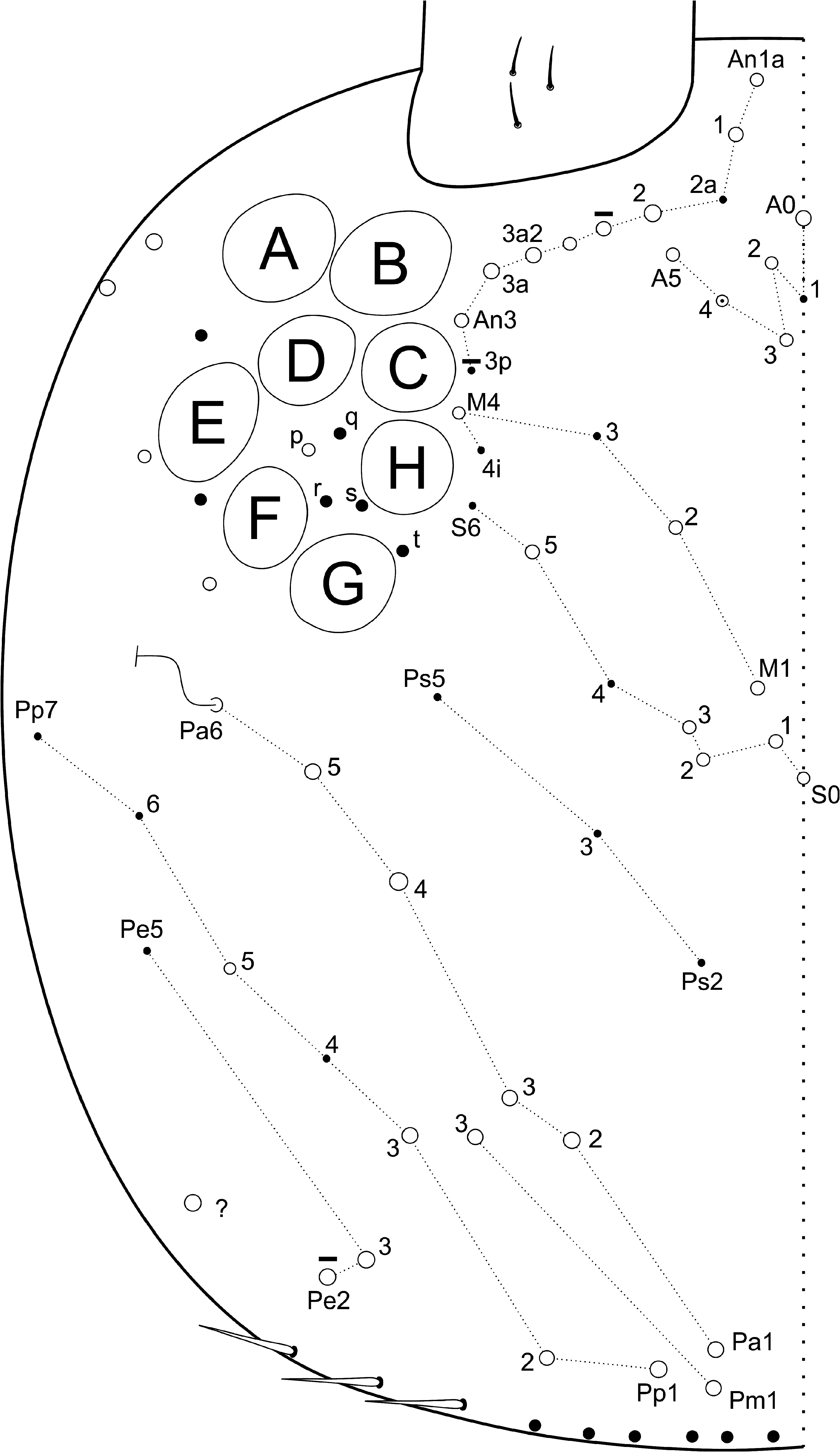
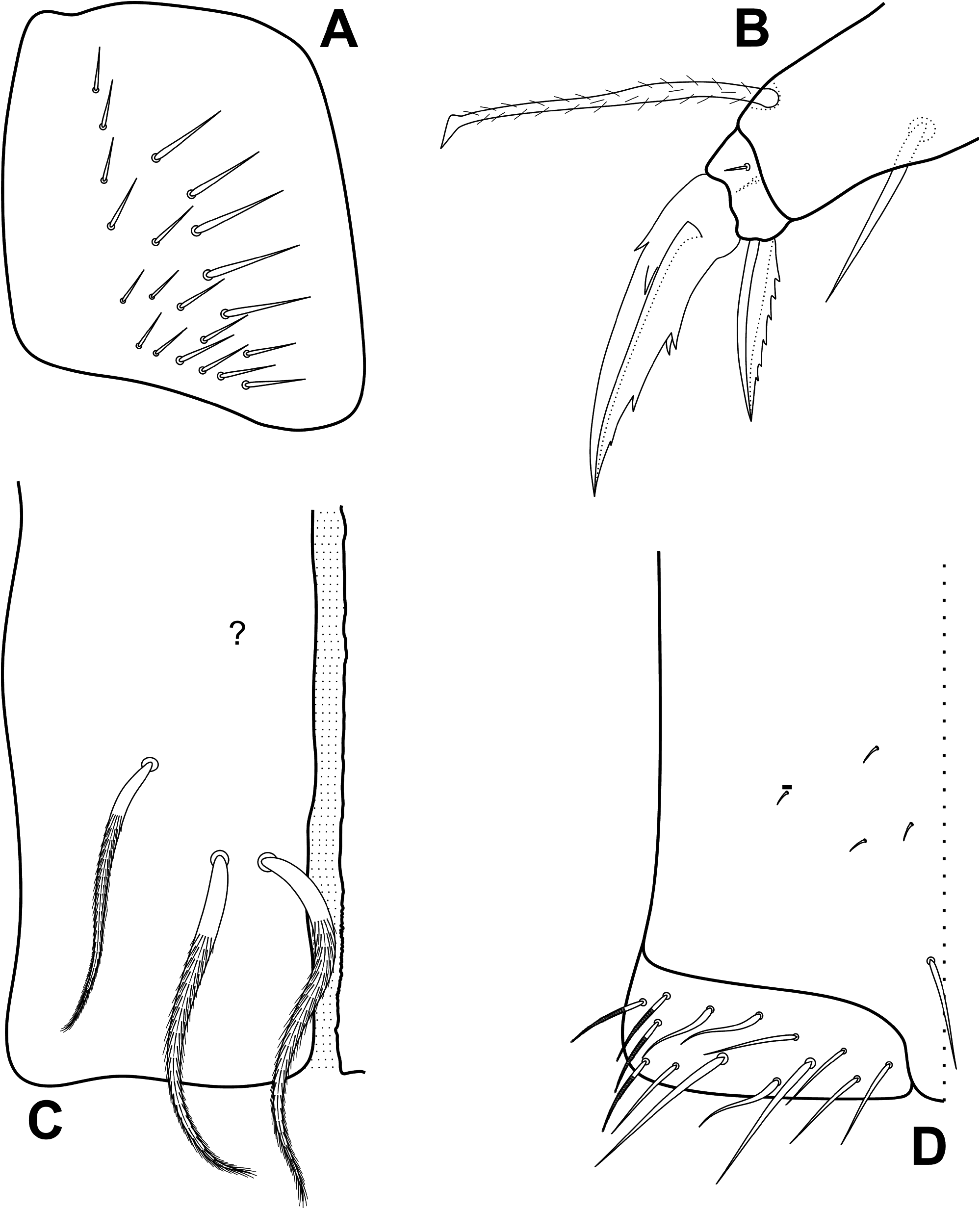
Head ( Figs 3–6 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 ): Ant. IV not annulated or subsegmented, with at least three types of chaetae: blunt sens, bristlelike sens and ciliated chaetae, apical bulb apically bilobed ( Fig. 3A View FIGURE 3 ). Ant. III sense organ with 2 rods, 2 small spiny and 1 blunt sens as guard-sensilla, plus some surrounding bristle-like sens, blunt sens and ciliated chaetae ( Fig. 3B View FIGURE 3 ). Ant. I dorsally with 3 sens-like mic at the base. Prelabral chaetae ciliated. Labral p0–1 chaetae longer than p2 and m series, a series shorter than m chaetae. Inner labral papillae conical, outer papillae almost imperceptible, as small protuberances. Labial basolateral and basomedian fields formula as M1–2rEL1–2a1–5, r chaeta reduced ( Fig. 4 View FIGURE 4 ). Labial palp with 5 proximal smooth chaetae, p3 and an2 smaller than the others ( Fig. 4 View FIGURE 4 ). Labial palp papillae and guard-chaetae formula as: H(2), A(0), B(5), C(0), D(4), E(4) + l.p. finger-shaped and surpassing the papilla base. Outer maxillary lobe with basal and distal chaetae subequal and smooth; sublobal plate with 3 smooth main appendages, basal one slightly smaller than the others, plus 1 outer reduced smooth appendage ( Fig. 4 View FIGURE 4 ). Ventral postlabial chaetotaxy as in Fig. 4 View FIGURE 4 , with: G1–4 ( G3 may be absent), H2–4, J1–4 anterior ciliated chaetae, with exception of J2 short and smooth, plus 2 other small smooth chaetae, posteriorly with 0–2 ciliated chaetae plus one b.c. ciliated ( Figs 4–5 View FIGURE 4 View FIGURE 5 ). Eye lenses A–B and E slightly larger than the others, other lenses subequal, with 5 interocular chaetae ( Fig. 6 View FIGURE 6 ). Dorsal macrochaetotaxy with 7–8 antennal ( An), 4–5 anterior ( A0, A2–5; A4 mac or mic), 3 medio-ocellar ( M1–2, M4), 5 sutural ( S0–3, S5), 5 post-occipital anterior ( Pa1–5), 2 post-occipital medial ( Pm1, Pm3), 4 postoccipital posterior ( Pp1–3, Pp5), 1–2 post-occipital external ( Pe2–3; Pe2 present or absent) plus one posterior mac without clear homology. Further details on the idiochaetotaxy are presented in Fig. 6 View FIGURE 6 .
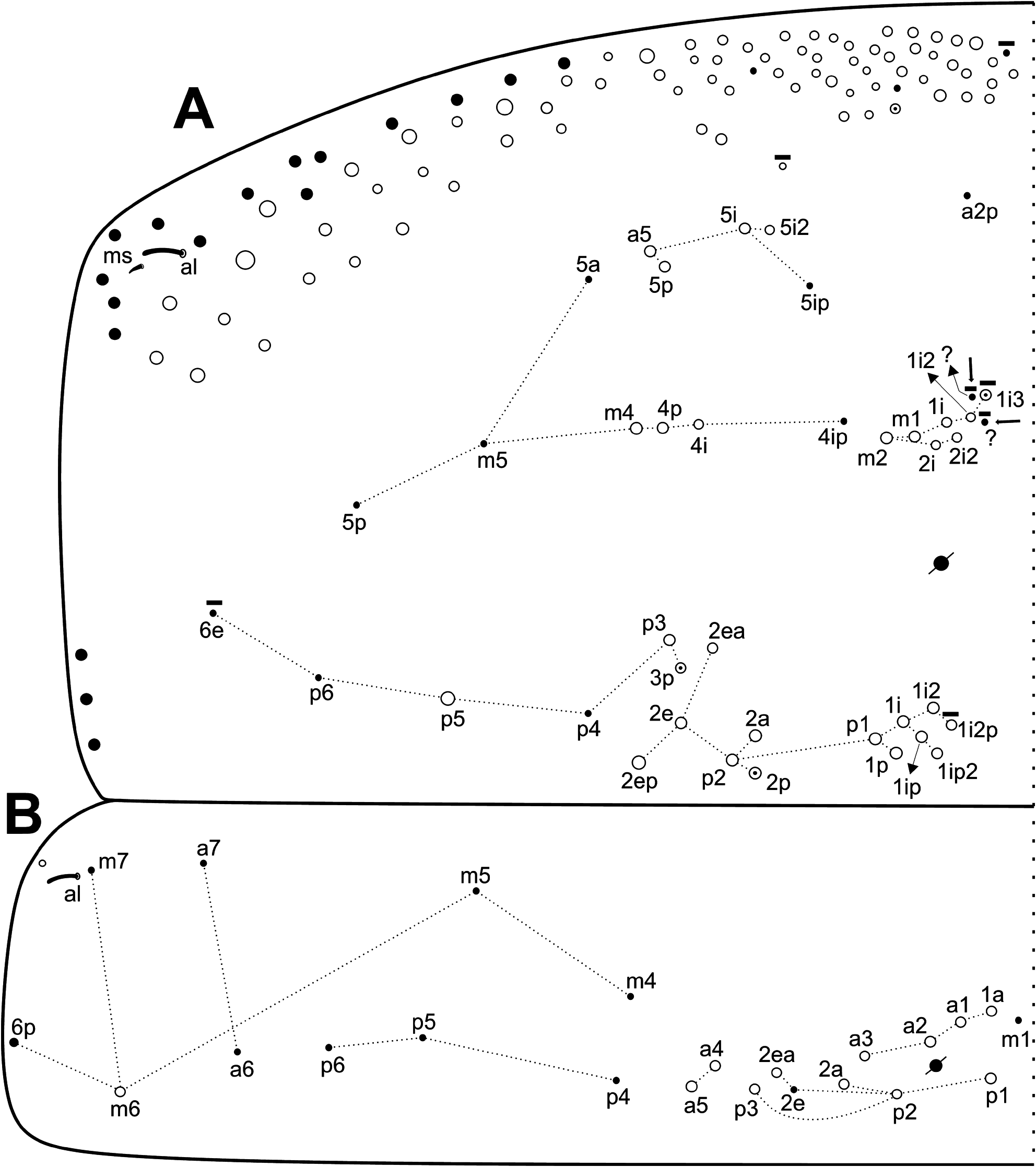
Trunk dorsal chaetotaxy ( Figs 7–9 View FIGURE 7 View FIGURE 8 View FIGURE 9 ): Th. II with 1 ms and 1 al; with 4 anterior ( a5–5p–5i2, excluding the anterior collar), 9–10 medial ( m1–1i3, m2–2i2, m4–4p–4i; m1i3 as mac or mic, present or absent) and 13–16 posterior ( p1–1p–1ip2–1i2p, p2–2a–2p, p2e–2ep–2ea, p3–3p, p5; p1i2p present or absent, p2p and p3p also as mic) mac
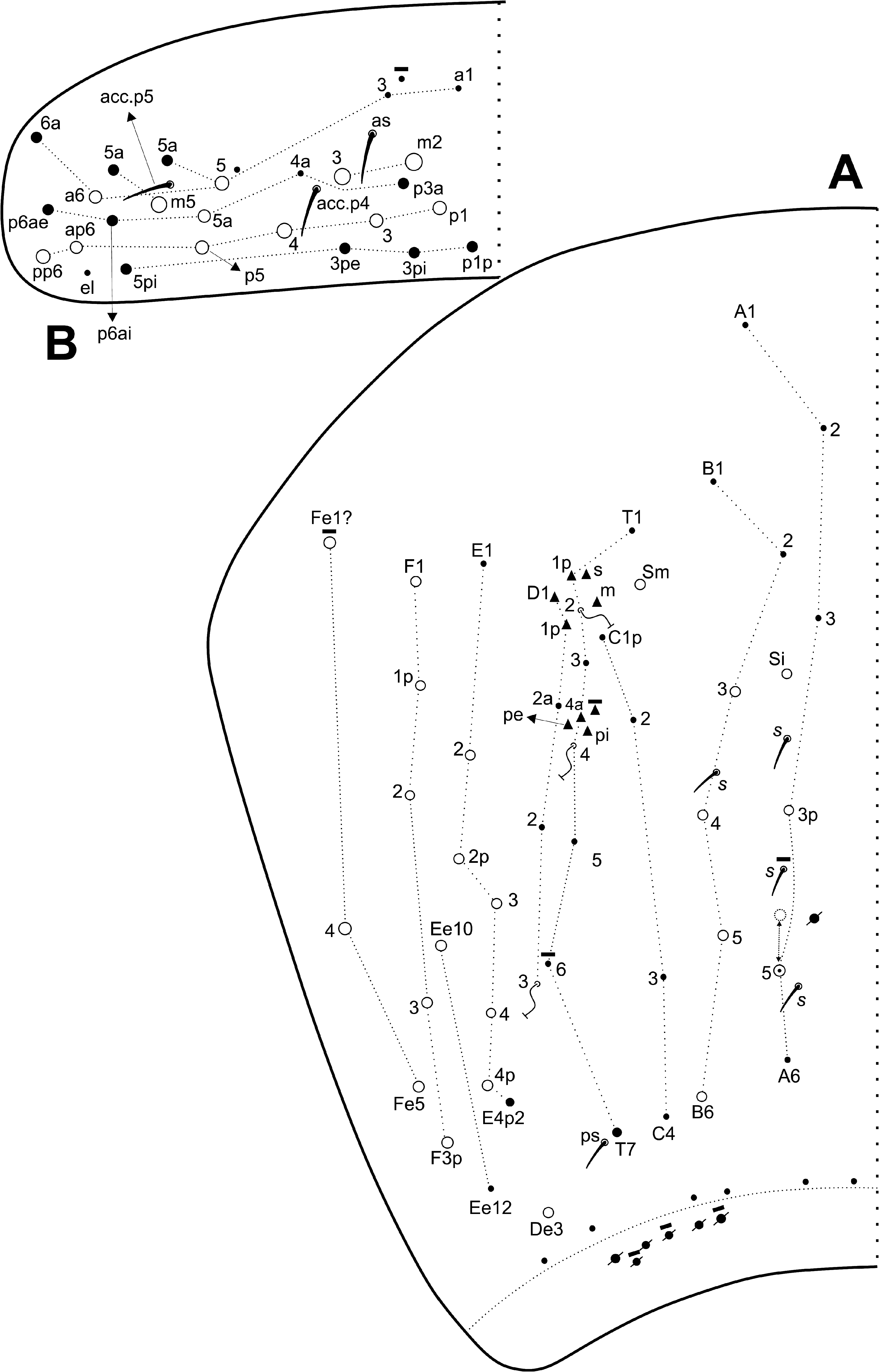
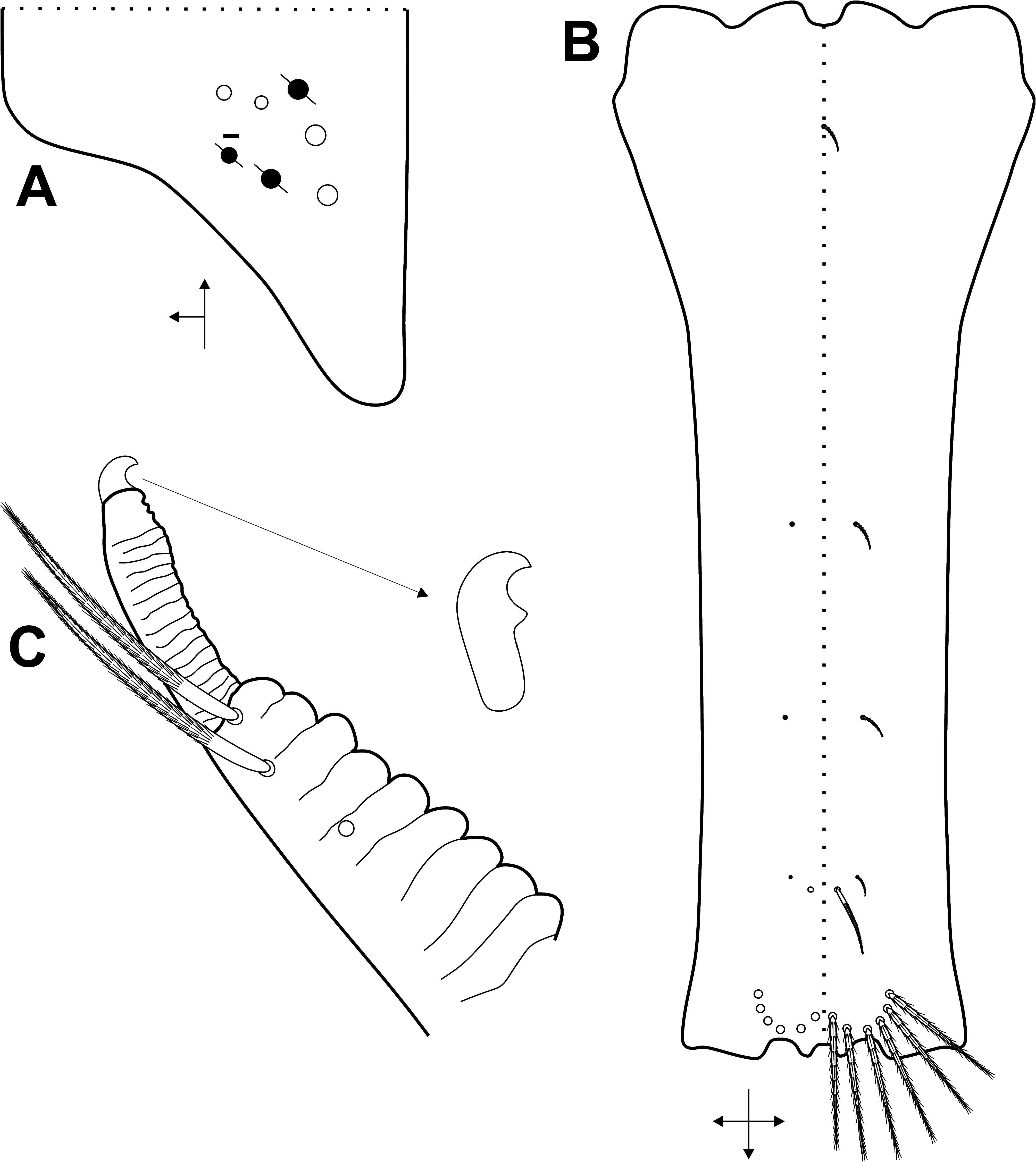
( Fig. 7A View FIGURE 7 ). Th. III with 1 al; 6 anterior ( a1–1a, a2–5), 1 medial ( m6) and 5 posterior ( p1, p2–2a–2ea, p3) mac ( Fig. 7B View FIGURE 7 ). Abd. I with 1 ms; and 4 medial ( m2–2i, m3, m4i) mac ( Fig. 8A View FIGURE 8 ). Abd. II with 1 as and 1 accessory sens ( acc. p6); 1 anterior ( a2) and 3 medial ( m3–3e, m5) mac, plus 2 bothriotricha ( a5, m2) ( Fig. 8B View FIGURE 8 ). Abd. III with 1 as, 1 ms and 1 accessory sens ( acc.p6), 3 medial ( m3, am6, pm6) and 3 posterior ( p6, p7–7i) mac, plus 3 bothriotricha ( a5, m2, m5) ( Fig. 8C View FIGURE 8 ). Abd. IV with at least 4–5 sens (possibly more), as apparently absent or not clearly distinguishable; mac formula as 1–2 ‘A’ ( A3p, A5; A5 as mac or mic), 4 ‘B’ ( B3–6), 1 ‘De’ ( De3), 5 ‘E’ ( E2–4p), 1 ‘Ee’ ( Ee10), 5 ‘F’ ( F1–3p), 2–3 ‘Fe’ ( Fe1?, Fe4–5; Fe1? may be absent), plus 2 secondary mac ( Si, Sm), 3 bothriotricha ( T2, T4, D3), and about 6 posterior mic and 3–6 psp ( Fig. 9A View FIGURE 9 ). Abd. V with 1 as and 2 accessory sens ( acc.p4–5); 2 anterior ( a5–6), 3 medial ( m2–3, m5), 1 posteroanterior ( p5a) and 6 posterior ( p1, p3–5, ap6–pp6) mac ( Fig. 9B View FIGURE 9 ). Further details on the idiochaetotaxy are presented in Figs 7–9 View FIGURE 7 View FIGURE 8 View FIGURE 9 .
Legs, collophore and furcula ( Figs 10–12 View FIGURE 10 View FIGURE 11 View FIGURE 12 ): Metatrochanteral organ with 12–22 spiny smooth chaetae (21 on holotype) ( Fig. 10A View FIGURE 10 ).Tibiotarsi not subdivided.Unguis with 4 inner teeth, 2 paired at the base, both equally sized, 1 median subequal to the basal and 1 smaller apical tooth; plus 1 pair of lateral teeth (only one side represented in the Fig. 10B View FIGURE 10 ) and 1 external unpaired tooth. Unguiculi lanceolate, with the postero-external lamella serrated. Tenent hair slightly ciliate and capitate.Tibiotarsus III with 1 inner distal smooth chaeta near the unguiculus( Fig. 10B View FIGURE 10 ).Ratio of the smooth chaeta: tenent hair: unguiculus: unguis of holotype as 1: 1.8: 1: 1.8. Collophore anterior side with 3 distal ciliated mac, other proximal ciliated chaetae unclear, with at least 3 proximal short spine-like chaetae ( Fig. 10C View FIGURE 10 , spine-like chaetae not represented); posterior side with 1 smooth chaetae plus 3–4 short spines; lateral flap with about 10 smooth and 4 ciliated chaetae, 2 smooth chaetae clearly longer than the others ( Fig. 10D View FIGURE 10 ). Manubrial plate with 4 ciliated chaetae, the internal 2 slightly larger than the lateral ones, and 2–3 psp ( Fig. 11A View FIGURE 11 ). Ventral manubrium with 1,2,2,2/2,12 central ciliated chaetae from the base to the apex ( Fig. 11B View FIGURE 11 ). Dorsal face of dens covered by ciliated chaetae of different sizes, forming two well defined rows that reach the distal portion, at the point where the dens becomes abruptly narrow ( Figs 11C View FIGURE 11 , 12 View FIGURE 12 ). Mucro typically falcate, with its proximal portion extending within the terminal projection of the dens cuticle, where it is possible to observe the marks of a smaller subapical tooth developed internally ( Fig. 11C View FIGURE 11 ).
Etymology. The new species was named after its type locality: Picos municipality, Piauí State, northeastern Brazil.
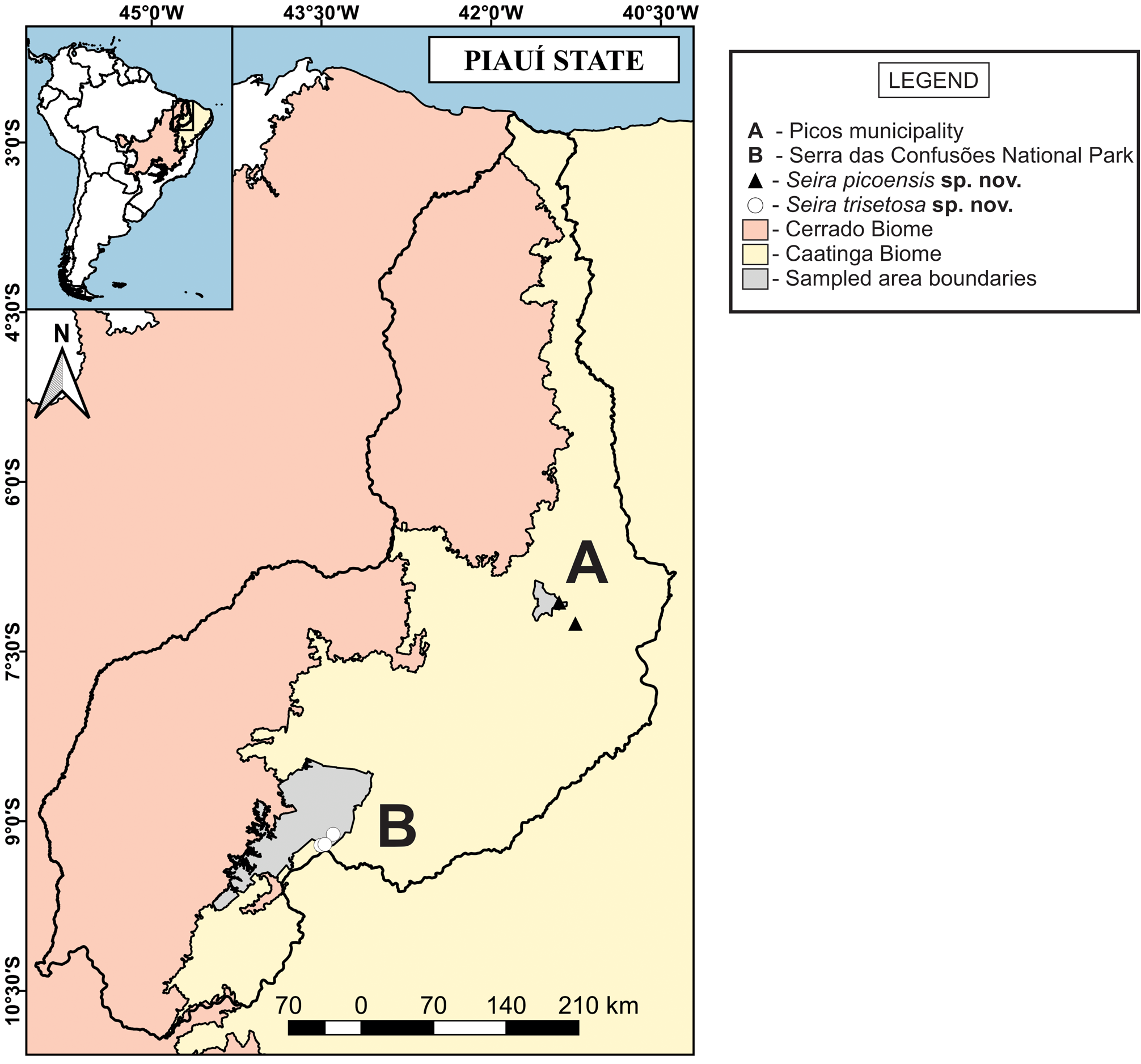
Distribution and habitat. The new species was found in two different localities: in the vicinities of IFPI (Instituto Federal de Educação, Ciência e Tecnologia do Piauí –Campus Picos), within the urban perimeter of the city; and in the surroundings of BR-407 highway, towards Jaicós municipality, Piauí State ( Fig. 13 View FIGURE 13 ). Both areas show clear signs of disturbance, with the presence of urban garbage, secondary vegetation in different degrees of ecological succession, recent signs of burning and proximity to highway heavy traffic. The region is inserted in the Caatinga domain and originally presents a typical deciduous vegetation, highly adapted to the semiarid climate, with a sandy soil poor in organic matter, with low water retention capacity and widely exposed to the sun. According to the Köppen-Geiger system, the climate of the area is “BShw”–Semiarid with a long dry season and rainy summer, characterized by high temperatures during almost all year and irregular precipitations ( Kottek et al. 2006). The annual average temperature is 27.9ºC, with maximum temperature reaching 39ºC and minimum 22ºC. The annual average rainfall is 679 mm, with the highest levels of precipitation from January to March ( CPRM 2004; INMET 2010).
Remarks. Among the Neotropical species of the genus, Seira picoensis sp. nov. resembles mostly S. bipunctata ( Packard, 1873) , S. caerucinerea Cipola & Bellini, 2014 (in Cipola et al. 2014a), S. delamarei Jacquemart, 1980b , S. dowlingi ( Wray, 1953) , S. fulva ( Schött, 1896) , S. manukio Soto-Adames, Bernard & Wynne (in Bernard et al. 2015), S. nicoya Christiansen & Bellinger, 1988 , S. socotrae Barra, 2004 , S. taeniata ( Handschin, 1925) and S. terrestris ( Folsom, 1932) , specially by the Abd. I with 4 mac ( S. terrestris with 2 or 4 mac) and Abd. II with 3 mac in A region ( S. caerucinerea and S. delamarei with 3–4 mac). The new species resembles only S. bipunctata , S. dowlingi and S. nicoya by labial palp papilla E l.p. morphology (finger-shaped); S. bipunctata , S. caerucinerea , S. delamarei , S. dowlingi , S. manukio , S. socotrae and S. terrestris by labial basomedian r chaeta reduced; S. bipunctata , S. caerucinerea , S. fulva , S. manukio , S. nicoya , S. taeniata and S. terrestris by head M2 chaeta as mac; S. caerucinerea , S. fulva , S. manukio , S. nicoya and S. taeniata by head M1 chaeta as mac; S. bipunctata , S. delamarei , S. dowlingi , S. fulva , S. manukio , S. nicoya , S. socotrae and S. taeniata by cephalic region 6 with 4 mac ( Pa2–3, Pm3, Pp3); S. manukio by head Pa4 chaeta as mac; S. bipunctata , S. dowlingi , S. manukio , S. nicoya and S. socotrae by head Pp5 chaeta as mac; S. bipunctata , S. caerucinerea , S. delamarei , S. dowlingi , S. manukio , S. socotrae , S. taeniata and S. terrestris by the unguiculus acuminate; and S. bipunctata , S. fulva , S. nicoya and S. terrestris by the presence of 2 manubrial subapical chaetae ( S. terrestris with 2–4). However, the new species is unique by the combination of: 1) labial palp papilla E with l.p. finger-shaped (pointed in S. caerucinerea and S. manukio ); 2) chaeta r of labial basomedian field reduced and smooth (normal and ciliated in S. fulva and S. nicoya ); 3) head M2 chaeta as mac (absent or as mic in S. delamarei , S. dowlingi and S. socotrae ); 4) head M1 chaeta as mac (absent or as mic in S. bipunctata , S. delamarei , S. dowlingi , S. socotrae and S. terrestris ); 5) head S4 chaeta as mic (mac in S. caerucinerea , S. fulva and S. nicoya ); 6) head S6i chaeta absent (as mac in S. caerucinerea ); 7) head S6 chaeta as mic (mac in S. bipunctata , S. caerucinerea , S. delamarei , S. dowlingi , S. manukio , S. socotrae and S. taeniata ); 8) head Pa4 chaeta as mac (absent or as mic in S. bipunctata , S. caerucinerea , S. delamarei , S. dowlingi , S. fulva , S. nicoya , S. socotrae and S. taeniata ); 9) head Pp5 chaeta as mac (absent or as mic in S. caerucinerea , S. fulva and S. taeniata ); 10) presence of 4 mac ( Pa2–3, Pm3 e Pp3) in the cephalic region 6 (as mic in S. caerucinerea ); 11) presence of 6–7 mac in the region 2 of Th. II (3 mac in S. fulva , 4 in S. bipunctata , S. delamarei , S. dowlingi , S. nicoya , S. socotrae and S. taeniata , and 4–5 in S. terrestris ); 12) presence of 6–7 mac in the region 3A of Th. II (5 mac in S. bipunctata , S. delamarei , S. dowlingi , S. fulva , S. nicoya and S. taeniata ); 13) presence of 11 mac in the regions A+B+C of Th. III (7 mac in S. nicoya , 8 in S. bipunctata , S. delamarei , S. dowlingi , S. fulva , S. socotrae and S. taeniata , 8–9 in S. terrestris , 9–10 in S. caerucinerea and 12 in S. manukio ); 14) presence of 7–8 central mac in Abd. IV ( 11 in S. caerucinerea and S. fulva , and 11–12 in S. nicoya ); 15) unguiculus acuminate (excavate in S. fulva and S. nicoya ); 16) anterior collophore chaetotaxy with at least 3 ciliated chaetae (1 chaeta in S. manukio ); 17) posterior collophore chaetotaxy with 1 smooth chaeta (2 ciliated chaetae in S. caerucinerea and S. nicoya , and 3 smooth chaetae in S. socotrae ); 18) manubrial plate with 4 ciliated chaetae (5 chaetae in S. dowlingi and S. socotrae , 6–7 in S. bipunctata and 7 chaetae in S. manukio ); and 19) ventral manubrium with 2 subapical chaetae ( 4 in S. caerucinerea , S. delamarei , S. dowlingi , S. manukio and S. socotrae ). Table 1 summarizes the morphological characteristics of Seira picoensis sp. nov. and of the compared species. In addition to the original descriptions, comparative information concerning S. bipunctata , S. delamarei , S. dowlingi , S. fulva and S. nicoya was extracted from Christiansen & Bellinger (2000); of S. bipunctata from Mari-Mutt (1987a); of S. dowlingi from Mari-Mutt (1986) and Soto-Adames (2008); of S. fulva from Christiansen & Bellinger (1988); of S. taeniata from Yoshii & Suhardjono (1989) and Yoshii (1990); and of S. terrestris from Christiansen & Bellinger (1992). Furthermore, specimens of S. dowlingi from Brazil, S. bipunctata and S. nicoya from Mexico were personally analysed.



| CC |
CSIRO Canberra Rhizobium Collection |
| R |
Departamento de Geologia, Universidad de Chile |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |