
Victorgorgia eminens, Moore & Alderslade & Miller, 2017
|
publication ID |
https://doi.org/10.11646/zootaxa.4304.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:3D557C94-0783-4C39-80C3-9C321DA94800 |
|
DOI |
https://doi.org/10.5281/zenodo.6015391 |
|
persistent identifier |
https://treatment.plazi.org/id/AF53AFBE-9F5A-4281-AC14-3473A66814FB |
|
taxon LSID |
lsid:zoobank.org:act:AF53AFBE-9F5A-4281-AC14-3473A66814FB |
|
treatment provided by |
Plazi |
|
scientific name |
Victorgorgia eminens |
| status |
sp. nov. |
Victorgorgia eminens sp. nov. Moore, Alderslade & Miller
http://zoobank.org/AF53AFBE-9F5A-4281-AC14-3473 A66814 View Materials FB ( Figs. 127–138 View FIGURE 127 View FIGURE 128 View FIGURE 129 View FIGURE 130 View FIGURE 131 View FIGURE 132 View FIGURE 133 View FIGURE 134 View FIGURE 135 View FIGURE 136 View FIGURE 137 View FIGURE 138 )
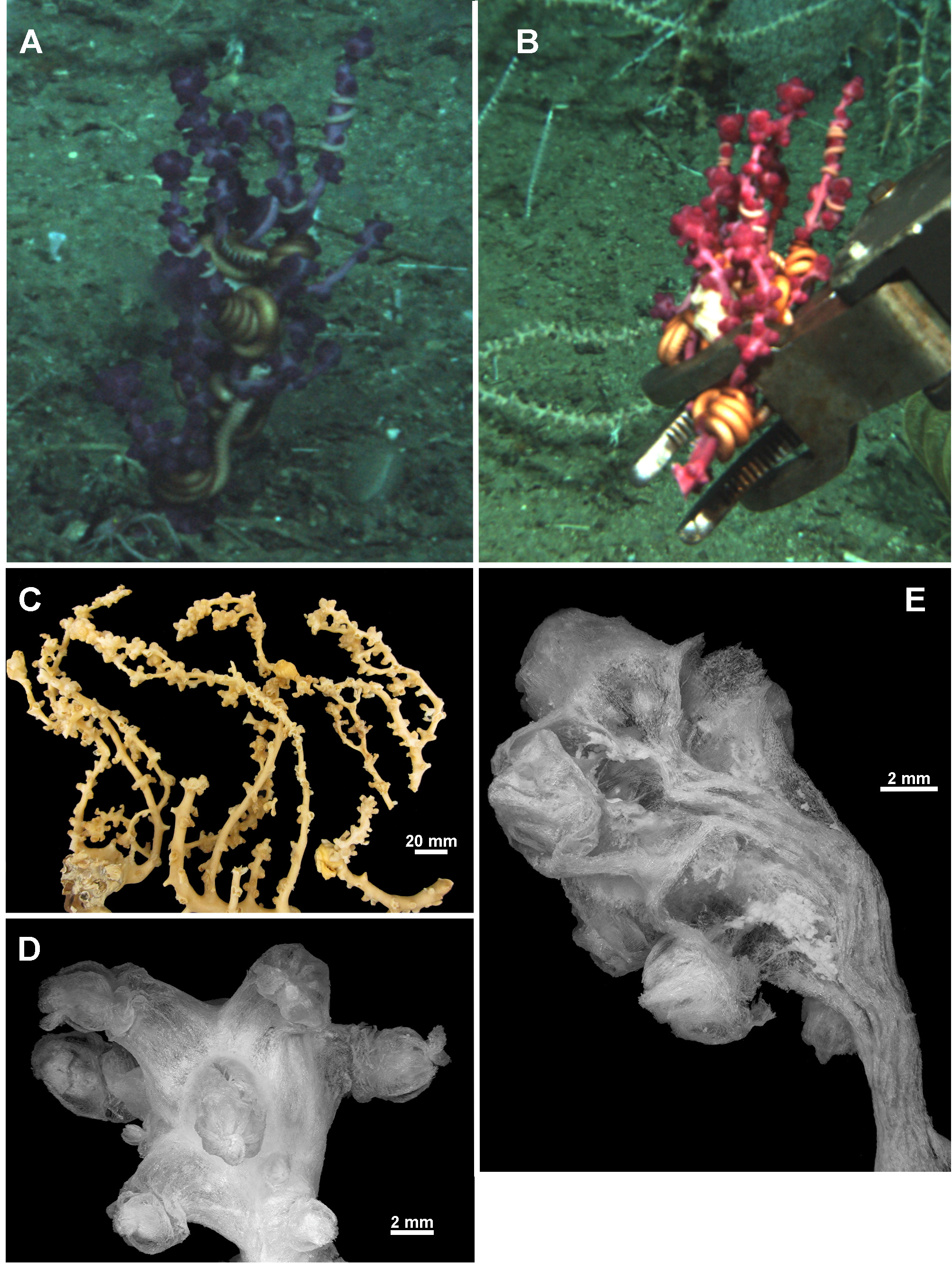
Material examined. Holotype: TMAG K4266, Z27 Seamount, Huon Commonwealth Marine Reserve (CMR), SW Tasman Sea, Australia, stn. J2-385-005, sample 010b, 44.245°S, 147.121°E, depth 1060 m, ROV Jason deployed from the U.S. RV Thomas T. Thompson, team led by Dr Jess Adkins & Dr Ron Thresher, 21st December 2008. GoogleMaps
Paratypes: TMAG K4267, Z27 Seamount , Huon CMR, SW Tasman Sea, Australia, stn. J2-385-005, sample 010a, 44.245°S, 147.121°E, depth 1060 m, ROV Jason deployed from the U.S. RV Thomas T. Thompson, team led by Dr Jess Adkins & Dr Ron Thresher, 21st December 2008 GoogleMaps ; TMAG K4268, Mongrel Seamount , Huon CMR, SW Tasman Sea, Australia, stn. J2-386-006, sample 0 0 1, 44.254°S, 147.115°E, depth 982 m, ROV Jason deployed from the U.S. RV Thomas T. Thompson, team led by Dr Jess Adkins & Dr Ron Thresher, 23rd December 2008 GoogleMaps ; TMAG K4269, Mongrel Seamount , Huon CMR, SW Tasman Sea, Australia, stn. J2-386-011, sample 0 21, 44.255°S, 147.114°E, depth 899 m, ROV Jason deployed from the U.S. RV Thomas T. Thompson, team led by Dr Jess Adkins & Dr Ron Thresher, 23rd December 2008 GoogleMaps ; TMAG K4270, Mongrel Seamount , Huon CMR, SW Tasman Sea, Australia, stn. J2-386-007, sample 0 0 3, 44.254°S, 147.114°E, depth 958 m, ROV Jason deployed from the U.S. RV Thomas T. Thompson, team led by Dr Jess Adkins & Dr Ron Thresher, 23rd December 2008 GoogleMaps ; TMAG K4115, Hill V Seamount , Huon CMR, SW Tasman Sea, Australia, CSIRO RV Southern Surveyor, stn. 69,(SS199701/69), 44.397– 44.398°S, 147.147– 147.178°E, depth 1262–1854 m, 31st January 1997 GoogleMaps ; NTM CO13052 (ex TMAG K1360), Hill J1 Seamount, SW Tasman Sea, Australia, CSIRO RV Southern Surveyor, stn. 36, (SS199701/36), 44.267– 44.242°S, 147.332– 147.363°E, depth 1518.4 m, 27th January 1997 GoogleMaps ; TMAG K4271, E.N.E. of St. Patricks Head , eastern Tasmania, Australia, CSIRO RV Soela, stn. 16, (SO198705/16), 41.573 S, 148.743 E, depth 1090–1150 m, 9th May 1987 GoogleMaps .
Other material: NTM CO13050 (ex TMAG K1362), Dory Hill Seamount, SW Tasman Sea, Australia, CSIRO RV Southern Surveyor, stn. 47, (SS199701/47), 44.322– 44.34°S, 147.115 –147.072°E, depth 1280–1400 m, 29th January 1997.
Description:
Colony form: The holotype is a large colony, about 230 mm high and 215 mm across, broken into two pieces, with an intact holdfast still attached to coral rubble ( Fig. 127 View FIGURE 127 A, B). The irregular, relatively sparse branching is principally in one plane. Basally, there is a main stem, with an approximate diameter of 8.6 mm, from which narrower branches emanate with a few instances of anastomoses with the holdfast. Higher in the colony, the stem becomes narrower with the diameter decreasing to 5.4 mm and side branch diameters between 3.4–5.3 mm in polyp-free areas. The branches are mostly circular in cross-section although tend to flatten at bifurcation points. Side branches occasionally branch again but are not crowded, with branches usually at least 15 mm apart. Anastomoses do not occur distally. The colony is in good condition although the tissues of the colony show evidence of dehydration from being frozen before being transferred to 70% alcohol.
Calyces are placed irregularly around the branches, generally at right angles, often bunched at points along the side branches and bunched tightly together on branch tips such that they form clavate clumps ( Fig. 127 View FIGURE 127 C, D). Frequently, there are parts of the branches without calyces, up to 45 mm long on the stem and 23 mm on the side branches. Occasionally there are isolated calyces, however more commonly they are crowded together in bunches with no or little space between them.
Colour: In the in situ photographs by the ROV Jason , the colony is deep purple, however the freshly collected specimen, when photographed before preservation, was closer to magenta ( Fig. 127 View FIGURE 127 B). The colony is now beige in alcohol.
Polyps and Calyces: The calyces are distinct, compact and firm when the polyps are retracted. Many polyps are partially retracted such that the base of the polyp head sits on the lip of the calyx and other polyps are fully retracted within the calyces which form 2–3.5 mm high, conical mounds with a small, round aperture ( Fig. 127 View FIGURE 127 D, E). The polyps and colony surface are covered with a thick, smooth integument which was partially removed to increase clarity in photographs ( Fig. 127 View FIGURE 127 Da, b). Fully exsert polyps are rare but can be up to 3.5 mm tall, measured from the lip of the calyx, although they more commonly extend only 2.5 mm. Usually the tentacles are folded over the mouth to form an eight-lobed, rounded polyp head which at the widest point is approximately 3.2–4.2 mm across; some juvenile polyps have a head diameter of approximately 2 mm. There is a single row of 8–10 pinnules along each side of the tentacles.
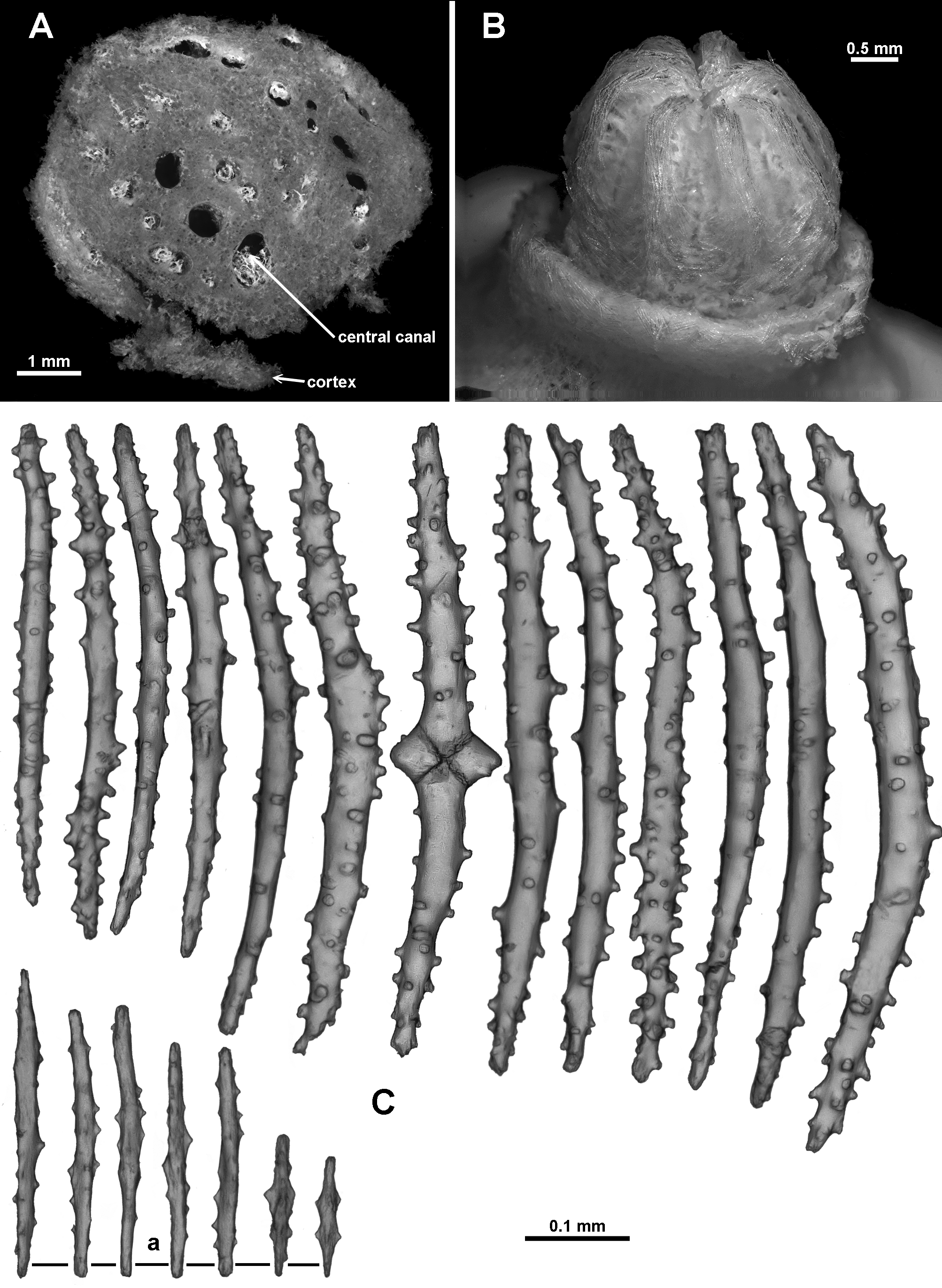
Medulla and Cortex: The branches of the colony consist of an essentially cylindrical medulla of tightly packed longitudinally arranged sclerites, surrounded by a thin cortex. Multiple adjoining longitudinal canals, which frequently anastomose to form a boundary space, separate the medulla from the cortex ( Fig. 127 View FIGURE 127 F). The few points of attachment between the cortex and medulla make it easy to dislodge the cortex. Two or three large coelenteric canals ( 0.7–0.9 mm diameter in the stem, 0.3–0.4 mm diameter in the peripheral branches) penetrate the medulla longitudinally and appear to extend throughout the colony ( Fig. 128 View FIGURE 128 A). Additional, smaller, indistinct canals occur adjacent to the larger canals.
The gastric cavities of the polyps on the branches terminate at the medulla and are connected by solenia extending through the cortex and into the boundary space. The body cavities of the polyps at branch tip extend some way down the branch and appear to coalesce with the central medulla canals.
Sclerites: Arrangements of the sclerites are largely obscured by the thick integument covering the colony. For examination this was removed by a short immersion in bleach and then washing in 70% ethanol. The polyp heads are covered in tightly packed tuberculate sclerites, arranged to form a collaret and points ( Fig. 128 View FIGURE 128 B). Transverse bunches of about 10 sclerites form the collaret at the base of the polyp head. These then grade en chevron up into the points, which continue longitudinally along the aboral side of the tentacles. The points and collaret sclerites are generally straight or curved, narrow sticks and spindles from 0.46–0.72 mm long which are sparsely covered in conical, flat-topped tubercles ( Fig. 128 View FIGURE 128 C). Amongst these occur smaller ( 0.11–0.29 mm), mostly smooth spindles with lateral, conical thorns ( Fig. 128 View FIGURE 128 Ca). Below the polyp head similar long, straight or slightly curved tuberculate sticks and spindles are arranged obliquely and sparsely on the polyp neck. On retraction of the polyp, these sclerites become more transverse and crowded and grade into the collaret. Most polyps on the holotype are wholly or partially retracted so the polyp neck is rarely visible.
On the aboral side of the tentacles, sclerites are commonly josephinae clubs with simple tubercles, more crowded on and near the clubbed end and absent or few on the handle of the club ( Fig. 129 View FIGURE 129 A). These crowded sclerites are 0.18–0.30mm long, decreasing in length distad, orientated with the clubbed end arranged towards the tentacle tip and bent upwards, making the aboral surface of tentacles prickly ( Fig. 129 View FIGURE 129 B). Densely arranged along the sides of the tentacle and projecting longitudinally into the pinnules are straight clubs or josephinae clubs, 0.2– 0.37mm long, with particularly narrow, mostly smooth handles ( Fig. 130 View FIGURE 130 ). Also crowded longitudinally in the pinnules are shorter, lightly tuberculate sticks and spindles and flat rods, 0.01–0.29mm long ( Fig. 130 View FIGURE 130 ), some with slightly clubbed tips.
No sclerites were found in the pharynx.
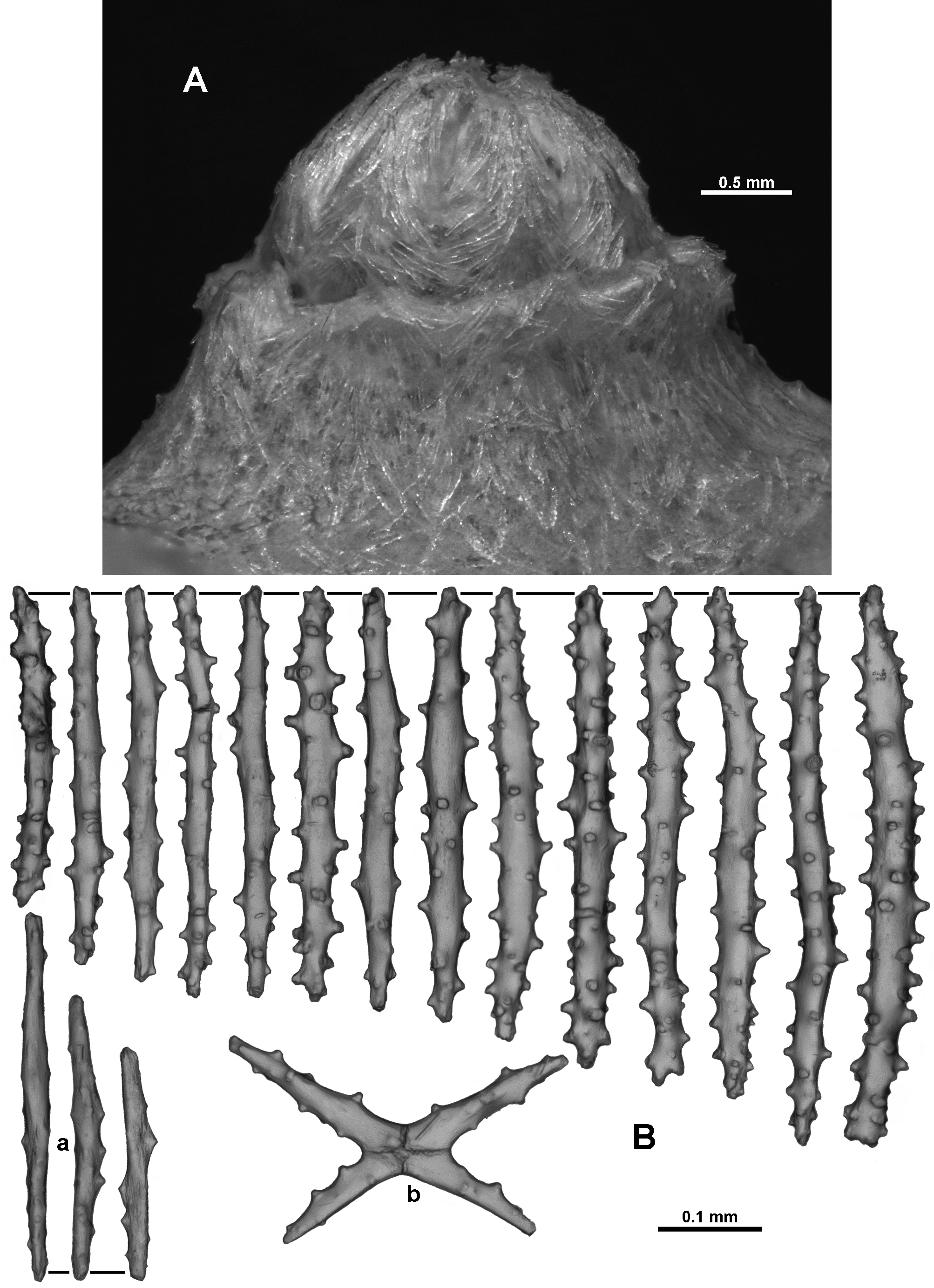
In the calyx, sclerites are arranged obliquely to longitudinally but grade to transverse at the base. They do not appear to form chevrons or peaks as the sclerites are arranged haphazardly ( Fig. 131 View FIGURE 131 A). They are mostly straight tuberculate sticks and spindles ( Fig. 131 View FIGURE 131 B), usually between 0.3–0.53mm long, with sparse, conical tubercles. Shorter, almost smooth, narrow spindles also commonly occur and range from 0.22–0.35mm long ( Fig. 131 View FIGURE 131 Ba). Very rarely, sparsely tuberculated crosses occur ( Fig. 131 View FIGURE 131 Bb).
The thin cortex commonly has straight tuberculate sticks and spindles ( 0.35–0.50 mm long) ( Fig. 132 View FIGURE 132 ), arranged longitudinally and obliquely on the stem and branches along with much more complex warty forms ( Fig. 132 View FIGURE 132 a) that occur patchily. In one particular sample the warty forms were more common than the tuberculate sticks and spindles but in other samples they were completely absent or rare. Most of them are between 0.14–0.35 mm long but some are up to 0.46 mm; they also tend to be wider than the smoother sticks and spindles. Occasionally, shorter, mostly smooth, narrow spindles similar to those from the calyx occur and, rarely, fused, cross or branched sclerites are present.
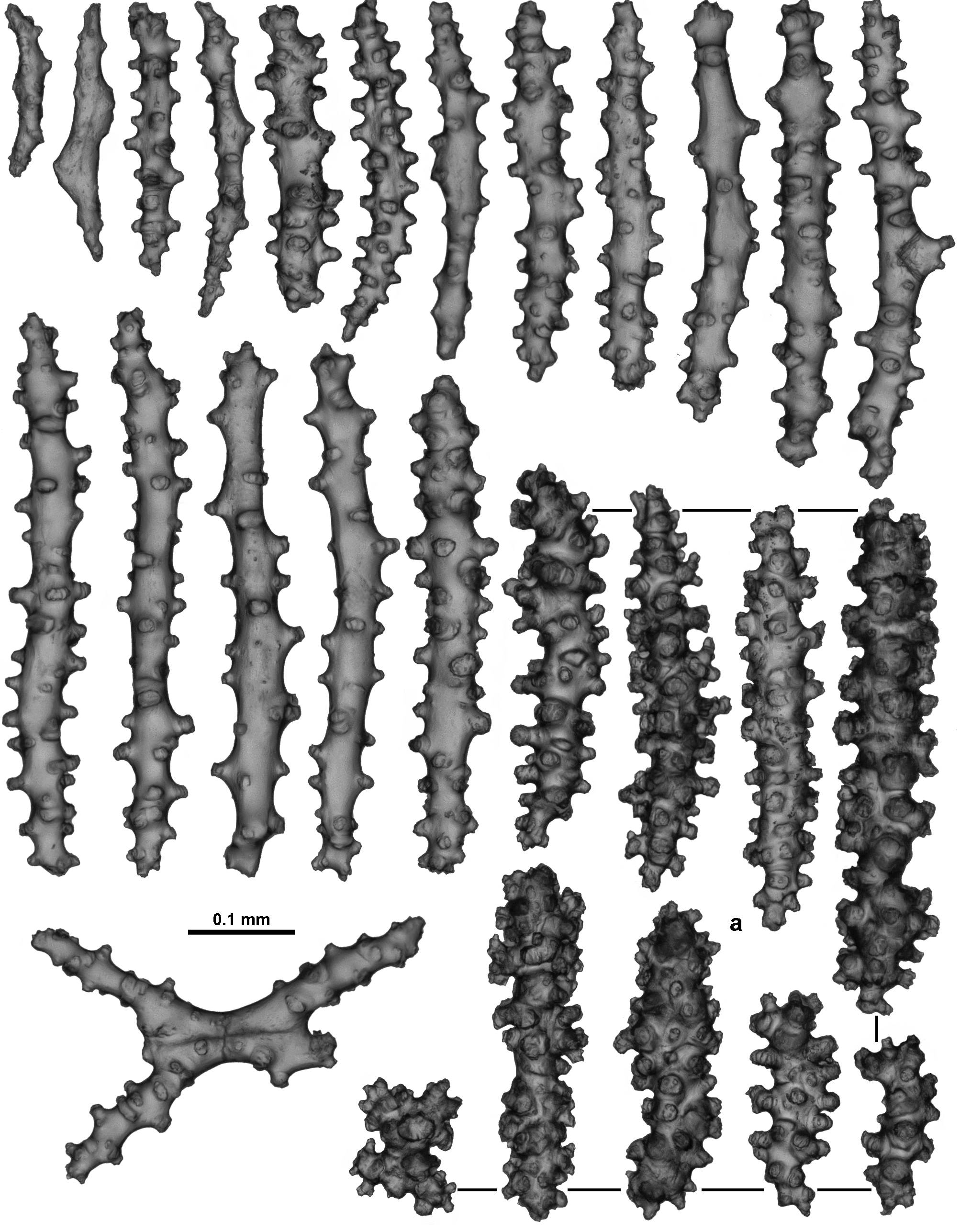
Tightly packed sclerites, mostly arranged longitudinally or obliquely, make up the medulla. These are mostly tuberculate or warty sticks and spindles, occasionally warty, along with some almost smooth, narrow forms ( Fig. 133 View FIGURE 133 ). Fused sclerites are occasionally present. Sizes range from 0.45–0.90 mm long.
Sclerites are uniformly transparent and colourless under transmitted light.
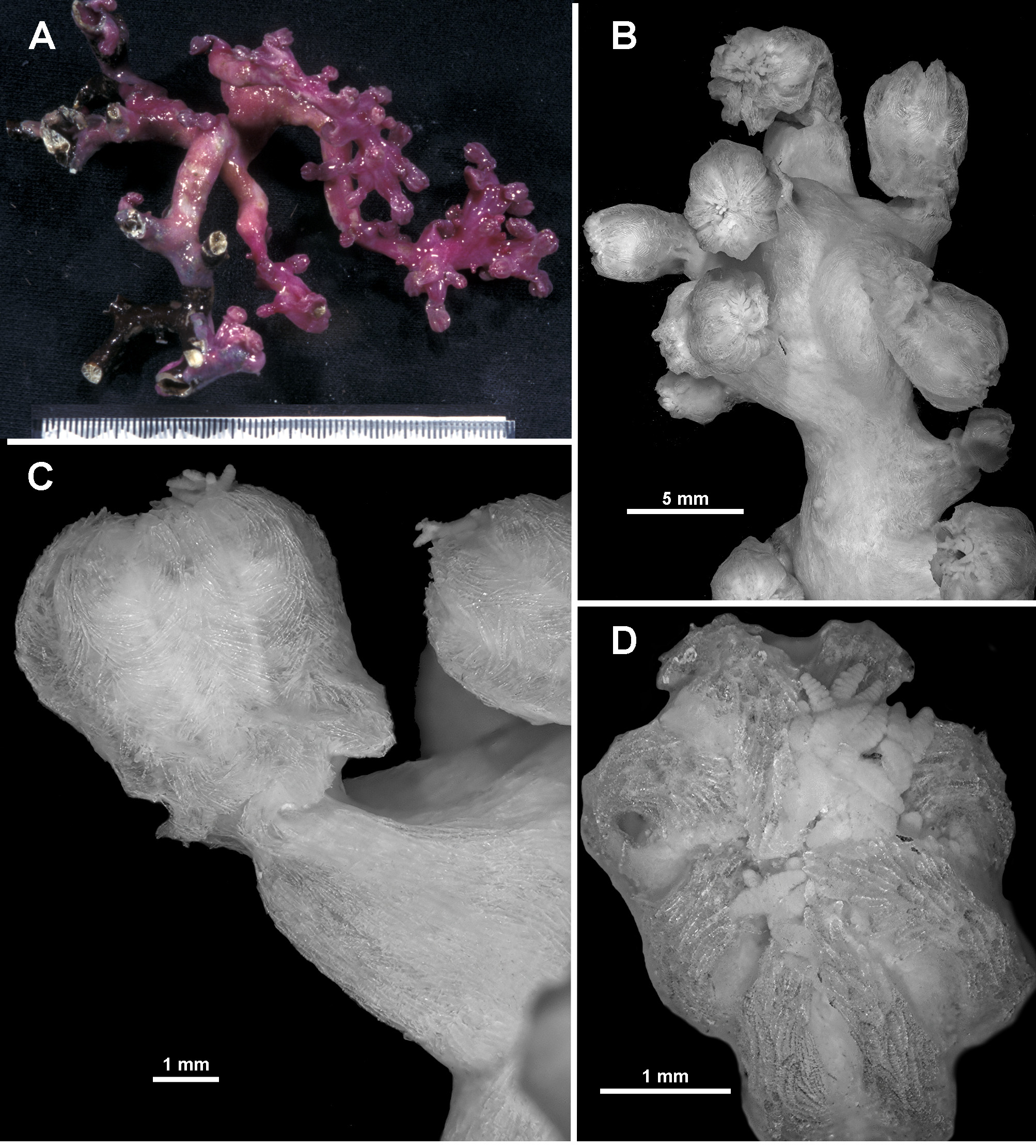
Variation: TMAG K4267 has a similar in situ colour to the holotype ( Fig. 134 View FIGURE 134 A) and is from a site very close to that from which the holotype was collected, however there are a few differences in colony form and sclerite ornamentation. A number of anastomoses evident amongst the colony branches and a small membranous holdfast encrusting coral rubble with at least two attachment points for the colony mean the colony form is more contorted than the holotype ( Fig. 134 View FIGURE 134 B). Sclerites from the tentacles are more complex than those from the holotype, particularly the handles of the clubs ( Fig. 134 View FIGURE 134 C), and the point sclerites are also more tuberculate. The specimen also has some calyx sclerites which are broader than those from the holotype, and tend to have a much more extensive cover of tubercles and sometimes complex warts ( Fig. 134 View FIGURE 134 D). No substantially warty sclerites were observed from the cortex. Other specimens examined also have calyx sclerites with slightly more complex warts and tubercles but these sclerites have patchy distributions and are not always present.
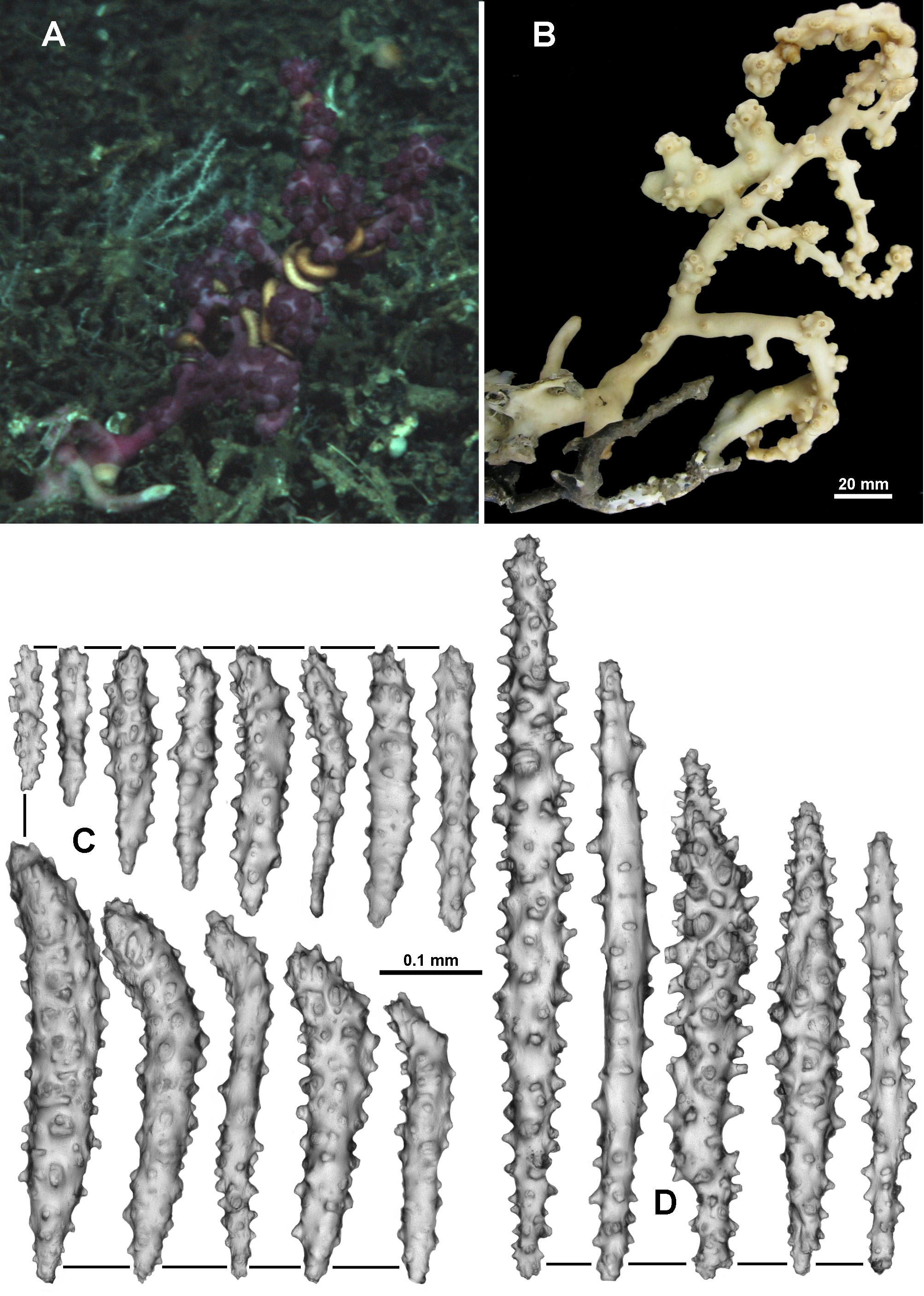
Another colony collected from a different seamount but still within the same area (TMAG K4268), was deep purple in one in situ photograph but magenta under closer lighting, like the holotype ( Fig. 135 View FIGURE 135 A, B). This colony has a branching pattern similar to the holotype, although not quite in one plane, with calyces clumped at branch tips and occasionally clumped along branches. However, in contrast to the holotype, calyces are also spread evenly and almost biserially, like V. josephinae , along the branches. Additionally, most polyps are fully exsert with almost none fully retracted, giving the preserved colony a slightly different appearance to that of the holotype.
Most of the other paratypes also have more exsert than retracted polyps giving the colonies a more untidy appearance than that of the holotype with its tightly retracted polyps. For example, TMAG K4271 is a large sample of many branched fragments ( Fig. 135 View FIGURE 135 C). The colony branches are narrower and floppier than the holotype with some long branches having few or no secondary branching. Calyces are spread widely along branches and very clearly grouped at branch tips such that the branches curl over from the weight of the bunched polyps. Most polyps are exsert and bent over ( Fig. 135 View FIGURE 135 D). A longitudinal cross-section of a terminal polyp bunch shows the polyp body cavities extending as canals down into the branch where they merge into the large coelenteric canals in the medulla ( Fig. 135 View FIGURE 135 E).
Another specimen, NTM CO13050, is only tentatively included in this species due to differences in the ornamentation and shape of the sclerites. The colony is a similar colour to the holotype ( Fig. 136 View FIGURE 136 A) but the polyps are larger and the calyces much more delicate than those of the holotype ( Fig. 136 View FIGURE 136 B, C). The sclerites in the tentacles have a similar arrangement to the holotype—josephinae clubs in the sides of the tentacles and longitudinally in the pinnules and clubs arranged along the tentacle rachis ( Figs. 136 View FIGURE 136 D; 137A), however, they lack the curved tip present in those of the holotype. The smaller tentacle rachis sclerites in particular tend to be straight, tuberculate rods, not true josephinae clubs ( Fig. 137 View FIGURE 137 Ba) while the larger sclerites are clubs that are more tuberculate than those in the holotype ( Fig. 137 View FIGURE 137 Bb). Additionally, in general, all sclerites are more tuberculate than the holotype, including those from the pinnules, calyx and cortex ( Fig. 138 View FIGURE 138 A–C).
Distribution: Southern seamounts and east coast of Tasmania, Australia
Depth: 899–1854 metres.
Remarks: V. josephinae has similar sclerite shapes and arrangements to V. eminens n. sp., however the most noticeable difference is the large josephinae clubs in the tentacles of V. josephinae in contrast to V. eminens n. sp., which has poorly developed josephinae clubs sometimes with slightly bent tips in the tentacles. Additionally, in V. josephinae the polyps tend to be arranged bi-serially while in this species there is a much greater tendency for the polyps to clump or bunch along and at the tip of the branches with notable regions of branches polyp-free.
Colour differences of the live colonies are striking—the colour of V. josephinae is recorded as “the coenenchyme ... was yellowish, while the anthocodiae were violet to deep purple” ( López-González & Briand 2002) in contrast to V. eminens n. sp. which is uniformly magenta to deep purple. However, colour is not always a reliable species characteristic and is not helpful for determination after preservation. Geographic distance between the two specimens is similarly noteworthy, as V. josephinae was collected off the coast of Portugal and V. eminens n. sp. off the coast of southern Australia.
The presence of large, dense, straight club sclerites in the points and aboral side of the tentacles of V. argentea n. comb. distinguishes it from V. eminens n. sp, while V. alba n. comb. and V. macrocalyx n. comb. both have bulky rods in the tentacle rachis with very few josephinae clubs, and V. nyahae n. sp. has many sharply pointed thorn clubs along the tentacle rachis, in the points, and in the calyx.
Etymology: The epithet is the participle of the Latin emino, eminent or prominent, in recognition of the fact that the large, purple specimens are very obvious and distinct in photographs and video footage.






No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
SubClass |
Octocorallia |
|
Order |
|
|
SubOrder |
Scleraxonia |
|
Family |
|
|
Genus |