
Valeriana officinalis, L. Valeriana officinalis subsp. hairy, L.
|
publication ID |
https://doi.org/10.1016/j.phytochem.2016.02.011 |
|
DOI |
https://doi.org/10.5281/zenodo.10530242 |
|
persistent identifier |
https://treatment.plazi.org/id/039ADC33-D904-FF89-7C6C-8ACDFABDFB7A |
|
treatment provided by |
Felipe |
|
scientific name |
Valeriana officinalisValeriana officinalis subsp. hairy |
| status |
|
2.4. Binary vector co-transformation of V. officinalis View in CoL hairy roots
Hairy root culture methodologies are common practice in many plant species, yet co-transformation with additional recombinant T-DNAs is comparatively underexplored as a means of metabolic engineering ( Chandra and Chandra, 2011). The Ri plasmid contains root-inducing loci (rol) A–D, which are essential for hairy root formation and growth ( Nilsson and Olsson, 1997), as well as the aux loci, which confers auxin autotrophy ( Nemoto et al., 2009). Rimediated transformation has been reported in species within the Valerianaceae clade ( Gränicher et al., 1992), however, no such protocols exist for binary vector co-transformation of this group. To engineer genes in addition to those mediating hairy roots, the binary vector pVo257 was introduced into A. rhizogenes harboring the Ri plasmid pRi15834. The T-DNA region of pVo257 contains a kanamycin selection marker plus a domain allowing for insertion of other genes of interest (GOI) flanked by the CaMV 35S promoter and a nopaline synthase terminator ( Fig. 6 View Fig ).
To establish conditions for the recovery of co-transformed lines, kanamycin sensitivity of hairy roots incorporating only the pRi15834 T-region was assayed. Approximately 100 mg root segments of a pRi15834 control line were inoculated onto solid MS medium containing increasing concentrations of kanamycin (0, 20, 40, 60, 80 mg /L). A dose-dependent reduction in growth was apparent across the range of concentrations tested. After 30 days of culture, new growth did not occur on media containing 80 mg /L kanamycin ( Fig. 7a View Fig ). Roots grown at this concentration of antibiotic were yellowed and the starting inoculum was otherwise unchanged. Thus, hairy roots transformed with A. rhizogenes harboring both pRi15834 and pVo257 plasmids were obtained by culturing explants on hormone-free MS medium (phenotypic selection for the pRi15834 T-Region conferring root differentiation and auxin autotrophic growth) followed by MS medium containing 80 mg /L kanamycin (selection for incorporation of the pVo257 T-DNA domain).
2.5. Overexpression of VoFPS and VoVDS genes in V. officinalis hairy roots
To more directly investigate the possible contribution of the VoFPS and VoVDS genes to overall sesquiterpenoid metabolism, co-transformed hairy root lines overexpressing these genes were compared to an empty vector control. The recombinant hairy root lines were initially selected for kanamycin resistance and subsequently confirmed for harboring the NPTII resistance gene and the respective targeted genes ( Fig. S4 View Fig ). Interesting morphological differences were noted in the respective cultures ( Fig. 8 View Fig ). The control line propagated via the development of thick taproots yielding proliferating lateral roots. In contrast, the VoFPS overexpression lines yielded less thickened roots. The VoVDS lines produced more of a labyrinth of thickened root phenotypes. While up to a 2-fold difference was noted in the development of biomass over a culture cycle of 7–10 days (control line growth> VoFPS overexpression lines> VoVDS overexpression lines), these growth differences have not been examined further.
As expected, the overexpression lines had increased transcript levels of the corresponding gene relative to the empty vector control line ( Fig. 9 View Fig ). The VoFPS lines featured 4–8-fold higher levels of VoFPS mRNA, which were not augmented further by MeJA treatment. The VoVDS overexpression lines featured 4–10-fold higher levels of the VoVDS mRNA, and these transcript levels were also not significantly affected by MeJA treatment. While the VoGCS mRNA level was not different in any of the recombinant lines relative to the empty control, the CYP71D442 mRNA was. Although not uniformly increased in all the VoFPS and VoVDS overexpression lines, CYP71D442 expression was increased almost 3-fold in several of the lines. However, this mRNA level was dramatically suppressed upon MeJA treatment just as it was in the empty control line.
To evaluate the efficacy of the binary vector co-transformation system for over-expression studies, an intron-containing β- glucuronidase gene inserted into the pVo257 vector was used as a reporter and histochemical marker. All of the kanamycin resistant lines exhibited positive GUS expression, but to varying extents. The examples shown in Fig. 7b View Fig represent relative high (GUS-1, GUS-3) and low (GUS-2) level expression. The degree of variable GUS staining could have as much to do with the permeability of the histochemical stain into root segments as expression variability in the various lines ( Fig. 7b View Fig ). As evident in Fig. 7c View Fig , uniform GUS gene expression was observed along the entire root axis, including individual root hairs of the co-transformed lines.
To examine how overexpression of VoFPS or VoVDS might influence carbon flux in these recombinant lines, the metabolic profile of each line was determined and compared to these same lines treated with 100 µM MeJA. Quantitation for select sesquiterpene components was extracted from the GC–MS chromatograms ( Figs. S5 View Fig and S 6 View Fig ). β- caryophyllene (5) and valerenadiene (6) were monitored as indicators of sesquiterpene hydrocarbon production, while valerenal (7) and valerenic acid (8) were monitored as indicators of carbon flux towards oxidized derivatives of valerenadiene. Compared to control levels, β- caryophyllene (5) content was increased in the VoFPS overexpression lines upwards of 3.5-fold ( Fig. 10a View Fig ). Both the VoFPS and VoVDS overexpression lines featured markedly increased valerenadiene (6) ( Fig. 10b View Fig ) and valerenal (7) ( Fig. 10c View Fig ). However, only the VoVDS lines accumulated increased valerenic acid (8) at levels 1.5- to 4-fold higher than control ( Fig. 10d View Fig ).
MeJA treatment stimulated sesquiterpenoid accumulation in all of the recombinant lines compared to untreated cultures. The highest valerenal (7) yields were achieved when overexpression lines were treated with MeJA ( Fig. 10c View Fig ). The highest valerenic acid (8) yields were achieved by MeJA treatment of the VoVDS overexpression lines ( Fig. 10d View Fig ). While some of the differences between the vector control line and the various engineered lines are modest and not statistically significant, the qualitative trends amongst the independent experimental lines are worth noting ( Figs. S5 View Fig and S 6 View Fig ). These findings suggest that VoFPS and VoVDS are promising targets to engineer V. officinalis roots for augmented sesquiterpenoid production. Synergistic elicitation of engineered lines for added production is a viable approach. However, the complex response to MeJA indicates that additional layers of regulation might be functioning in V. officinalis roots and further investigation is necessary to understand the signal transduction leading to metabolite production.
Chandra, S., Chandra, R., 2011. Engineering secondary metabolite production in hairy roots. Phytochem. Rev. 10, 371 - 395. http: // dx. doi. org / 10.1007 / s 11101 - 011 - 9210 - 8.
Granicher, F., Christen, P., Kapetanidis, I., 1992. High-yield production of valepotriates by hairy root cultures of Valeriana officinalis L. var. sambucifolia Mikan. Plant Cell Rep. 11, 339 - 342. http: // dx. doi. org / 10.1007 / BF 00233361.
Nemoto, K., Hara, M., Goto, S., Kasai, K., Seki, H., Suzuki, M., Oka, A., Muranaka, T., Mano, Y., 2009. The aux 1 gene of the Ri plasmid is sufficient to confer auxin autotrophy in tobacco BY- 2 cells. J. Plant Physiol. 166, 729 - 738. http: // dx. doi. org / 10.1016 / j. jplph. 2008.09.006.
Nilsson, O., Olsson, O., 1997. Getting to the root: the role of the Agrobacterium rhizogenes rol genes in the formation of hairy roots. Physiol. Plant. 100, 463 - 473. http: // dx. doi. org / 10.1111 / j. 1399 - 3054.1997. tb 03050. x.
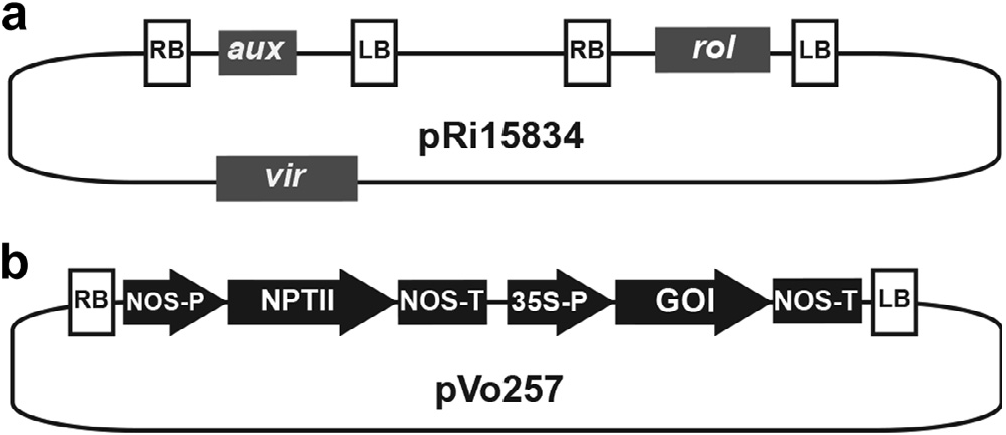
Fig. 6. Schematic diagrams of the plasmids used for A. rhizogenes-mediated binary vector co-transformation of V. officinalis hairy roots. (a) A. rhizogenes root-inducing plasmid pRi15834. The T-region comprises the aux and rol loci, which occur on separate T-DNAs. Transfer and integration of the T-region is mediated by the vir locus, whose gene products act in trans on 25-bp border repeat sequences to the right (RB) and left (LB) of T-DNA domains.(b) pBin derived binary vector Vo257.The T-DNA contains the selection marker neomycin phosphotranferase (NPTII), which is flanked by the nopaline synthase promoter (NOS-P) and nopaline synthase terminator (NOS-T). The CaMV35s promoter (35S-P) drives constitutive expression of the downstream gene of interest (GOI).
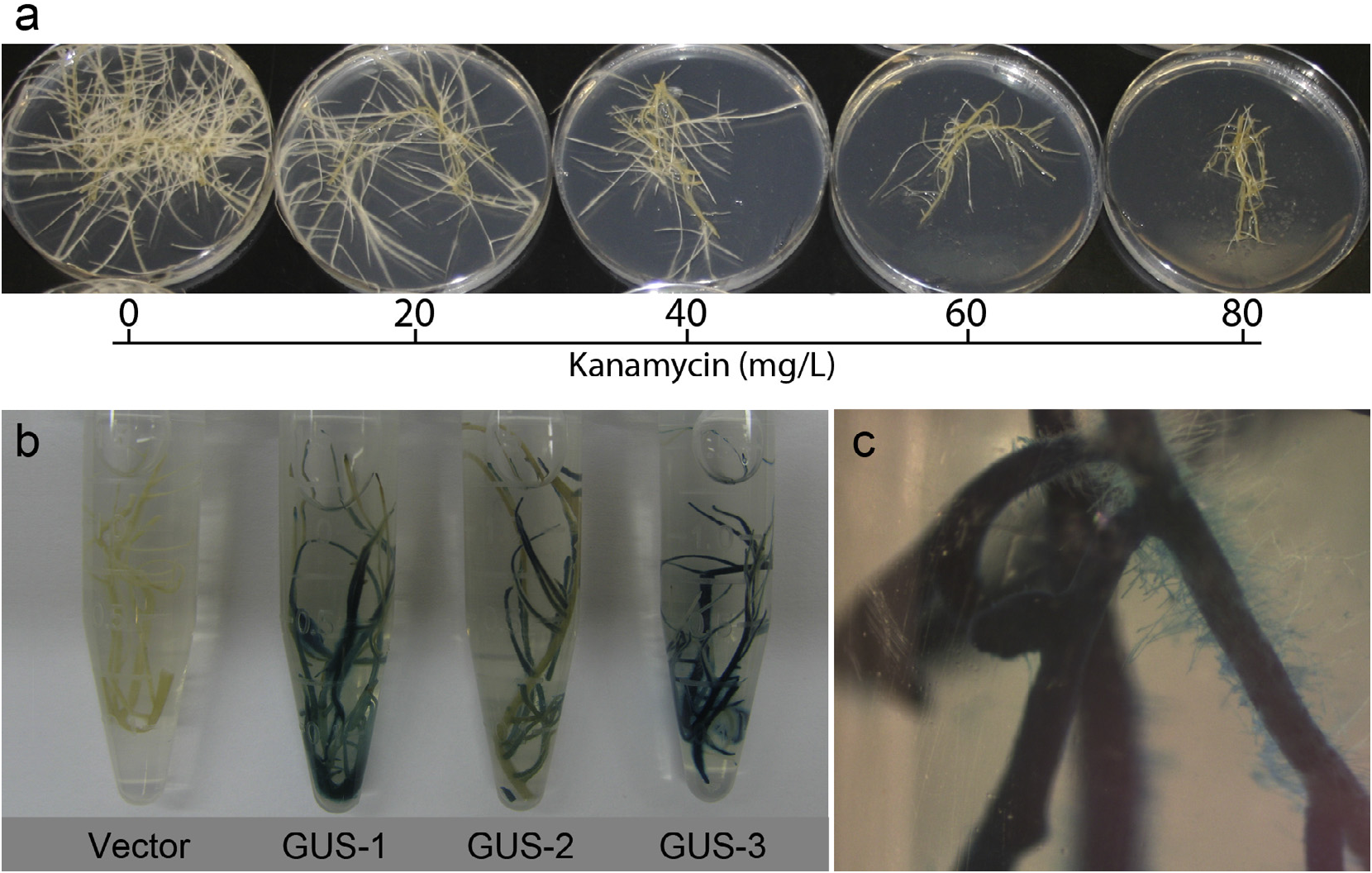
Fig. 7. Establishment of binary vector co-transformation conditions and heterologous gene expression. (a) Kanamycin sensitivity assay of pRi15834 control hairy roots. Root tips were inoculated to MS medium containing a range of kanamycin concentrations (0, 20, 40, 60, 80 mg/L) and qualified after 30 days. (b) GUS histochemical staining of cotransformed hairy roots. The lines harbored the pVo257 T-DNA, which contained a GUS-intron overexpression cassette, or the empty vector control (pVo257 EV). The three independent GUS lines exhibited varying degrees of staining, whereas no staining occurred in the empty vector line (C) Magnified visualization of GUS expression along the root axis including individual root hairs.
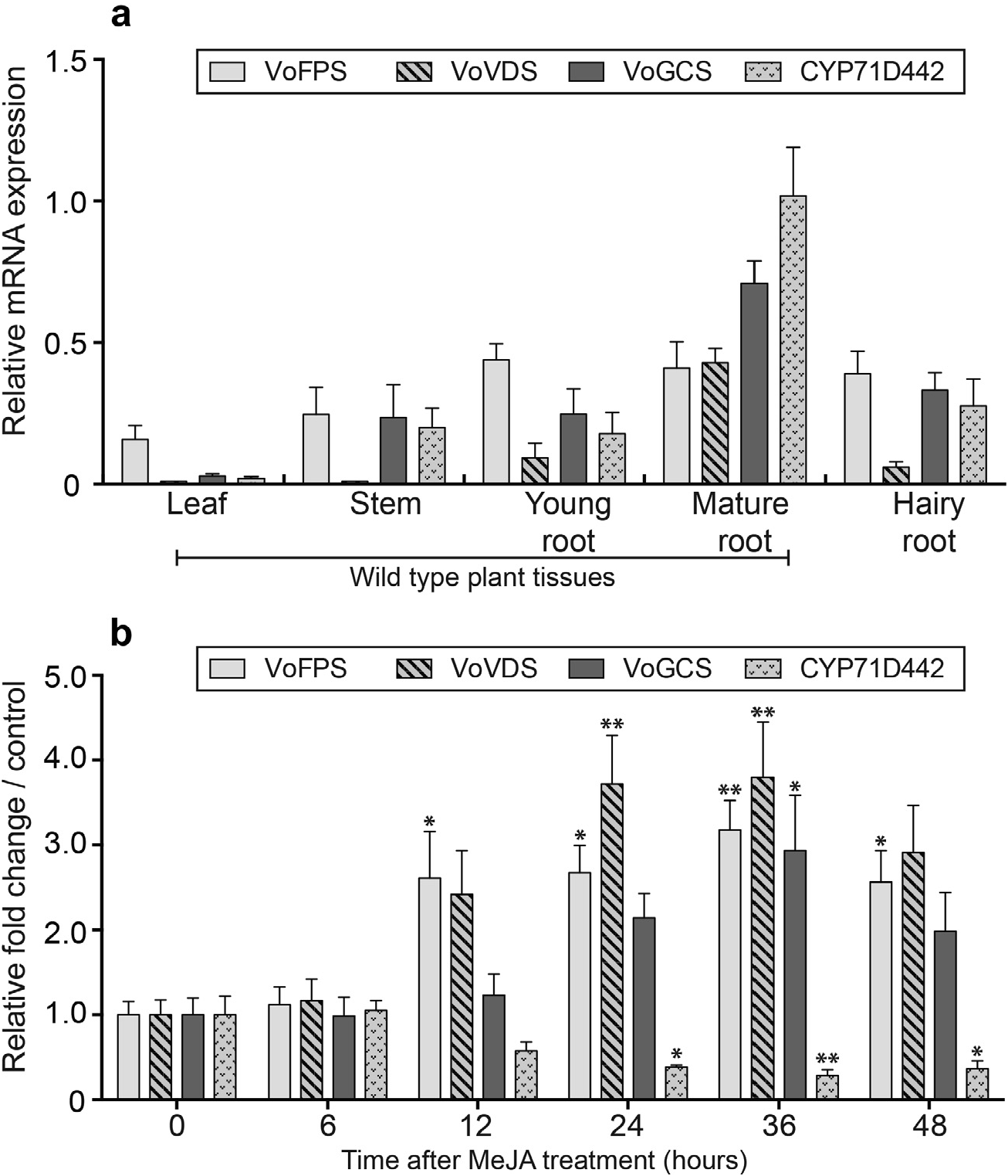
Fig. 4. Quantitati-ve real time polymerase chain reaction (qRT-PCR) expression profiling of VoFPS, VoVDS, VoGCS and CYP71D442 transcripts in wild-type V. officinalis plant tissues and hairy roots. (a) Relative transcript levels in leaf, stem, young root, mature root wild-type tissues and hairy root. (b) Time course of transcript levels in methyl jasmonate (MeJA) treated hairy root cultures. Actin expression levels were used as a reference for normalization. Three biological and technical replicates were used to certify the accuracy of the results and to calculate the standard error for each sample. For time course, asterisks indicate statistical significance in comparison to 0 h control assessed by one-way ANOVA (**, P <0.01; *, P <0.05).
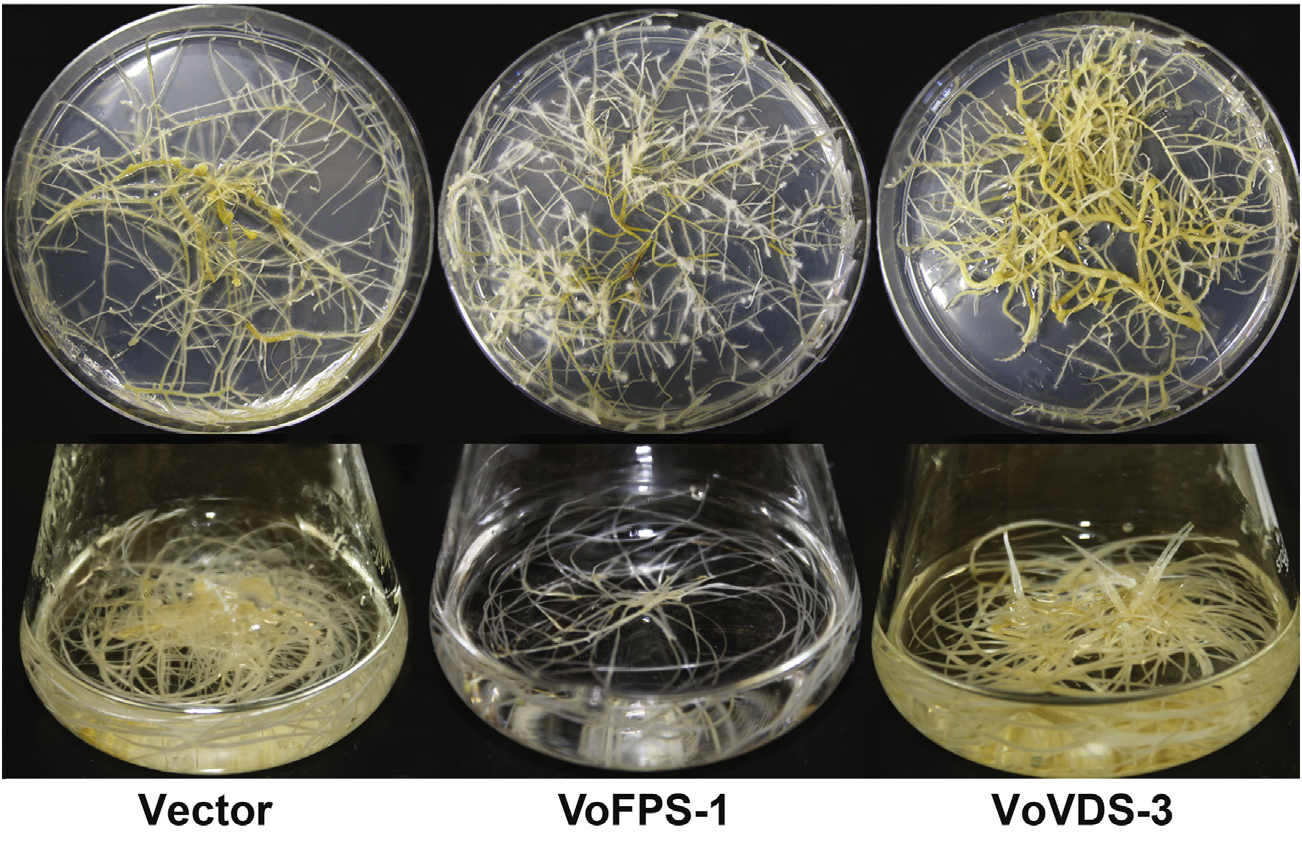
Fig. 8. Representative phenotypes of binary vector co-transformed V. officinalis hairy root lines grown on solid media and in liquid cultures. Showing empty vector control (pVo257 EV) compared to lines generated for overexpression of the VoFPS (VoFPS-1) and VoVDS (VoVDS-3) genes.
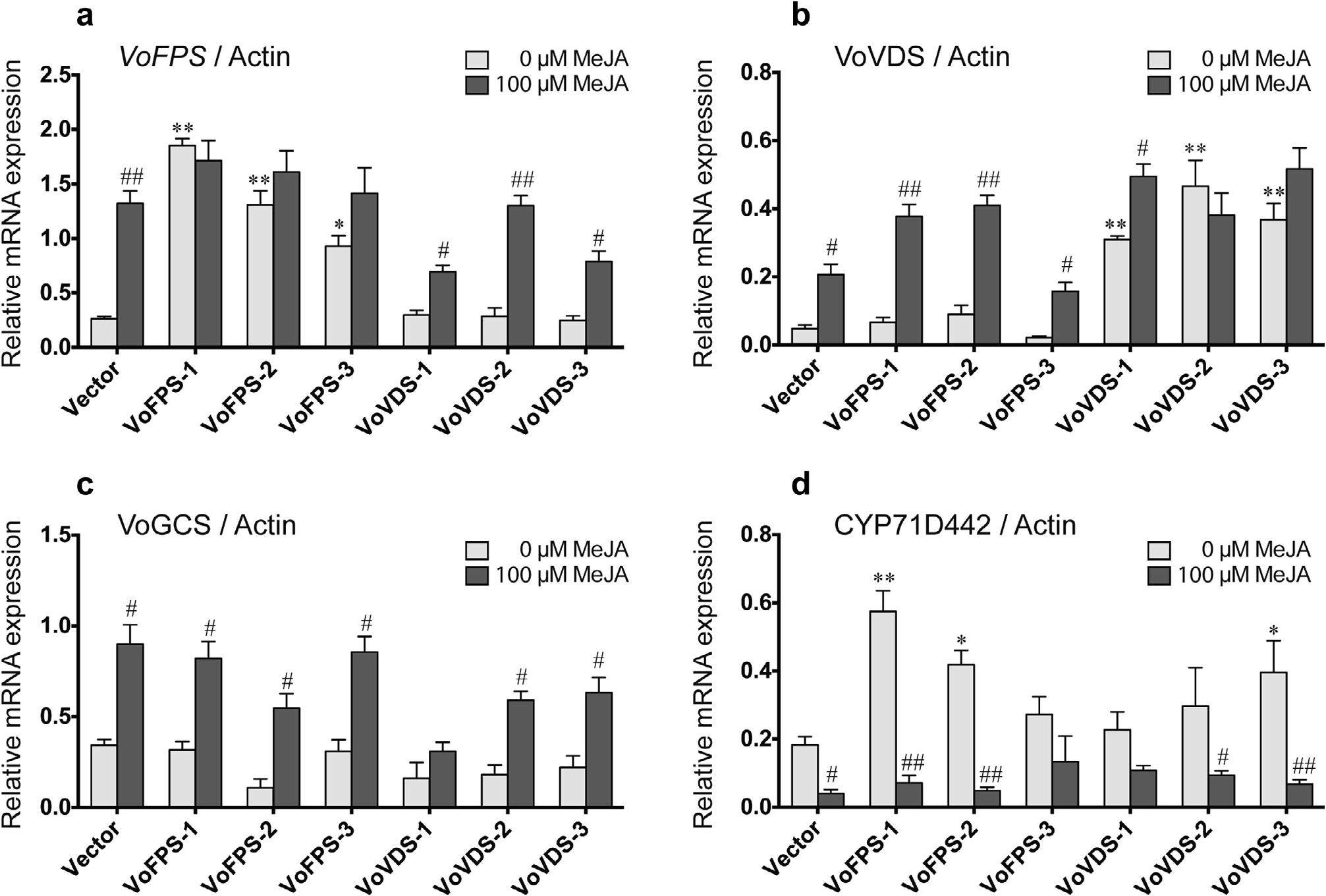
Fig. 9. Transcriptional profiling of recombinant V.officinalis hairy root lines including vector control and three independent lines overexpressing the VoFPS gene (lines 1–3) or the VoVDS gene (lines 1–3). qRT-PCR was performed following treatment with 0 µM or 100 µM MeJA for 36 h. Actin expression levels were used as a reference for normalization. Three biological and technical replicates were used to certify the accuracy of the results and to calculate the standard error for each sample. Asterisks indicate statistical significance in comparison to empty vector control assessed by one-way ANOVA (**, P <0.01; *, P <0.05). Hashes indicate statistical significance between MeJA treatments for each line as evaluated by students t-test (##, P <0.01; #, P <0.05).
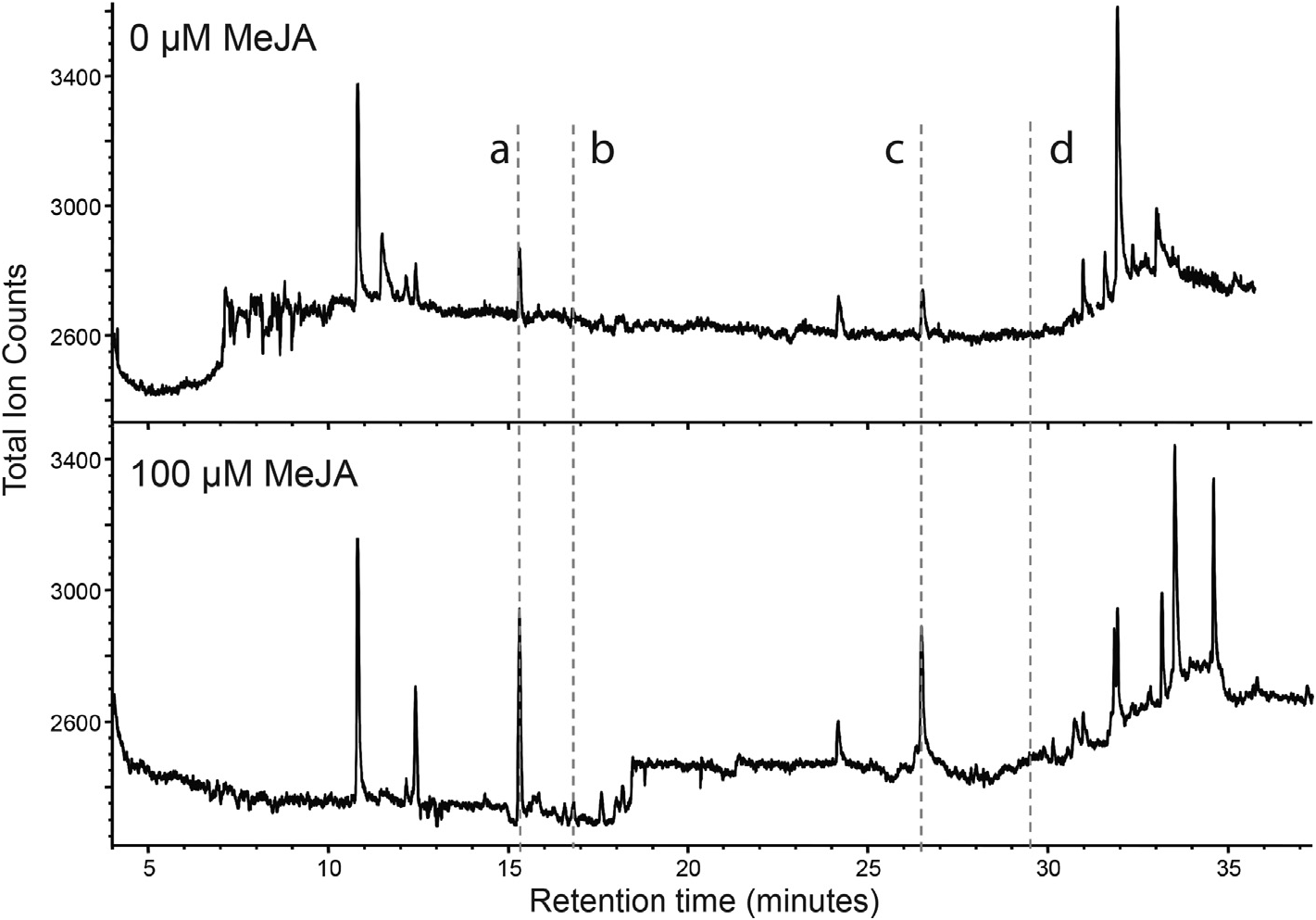
Fig. 5. GC–MS profile comparisons of sesquiterpenoid chemistry of hairy root cultures treated with methyl jasmonate (MeJA). Isogenic cultures were treated with 0 µM (control) (panels A, B) or 100 µM MeJA (panels C, D) and harvested for chemical analysis after 36 h. Hexane extracts prepared from the indicated cultures were analyzed directly (A, C) and after derivatization (B, D) by GC–MS. The peaks for β-caryophyllene (a), valerenadiene (b), valerenal (c) and valerenic acid (d) are annotated to facilitate comparisons.
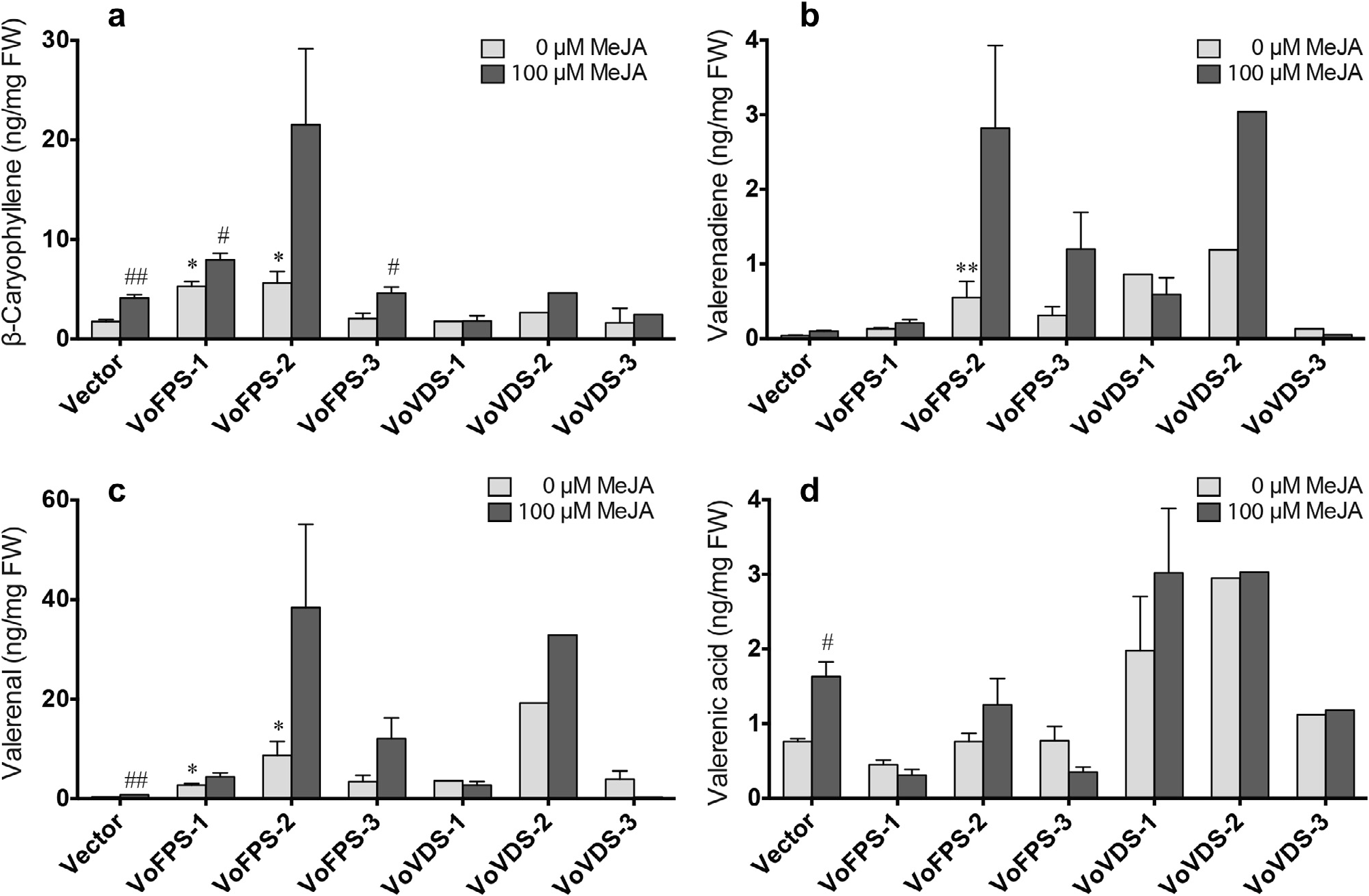
Fig. 10. Quantification of sesquiterpenoids in recombinant V. officinalis hairy root lines, including vector control and three independent lines overexpressing the VoFPS (lines 1–3) or VoVDS (lines 1–3) genes. Cultures were treated with 0 µM or 100 µM MeJA for 36 h and hexane extracts were prepared from the indicated lines from at least two samples and profiled by GC–MS prior to quantification of the indicated sesquiterpenoid relative to an internal standard.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |