
Rhinella yanachaga, Lehr, Edgar, Pramuk, Jennifer B., Hedges, Blair & Córdova, Jesús H., 2007
|
publication ID |
https://doi.org/ 10.5281/zenodo.179969 |
|
DOI |
https://doi.org/10.5281/zenodo.5625810 |
|
persistent identifier |
https://treatment.plazi.org/id/039A87E6-023A-B33F-CA8B-F9E38071F95D |
|
treatment provided by |
Plazi |
|
scientific name |
Rhinella yanachaga |
| status |
sp. nov. |
Rhinella yanachaga View in CoL new species
Holotype: MHNSM 19994 ( Fig. 1 View FIGURE 1 ), an adult male from Provincia de Pasco, Departamento de Pasco, National Park Yanachaga-Chemillén, W side of the Cordillera Yanachaga near Río San Alberto (= 2.9 km N, 5.5 km E [airline] Oxapampa), ca. 1 km N, 14 km E. Oxapampa by road on trail, 2600 m elevation, Peru, obtained by J. Icochea on an unknown date in the late 1990s.
Paratypes: 29, all from Provincia de Pasco, Departamento de Pasco, Cordillera de Yanachaga , Peru: 14 females ( MHNSM 24510–17, 24519–22, MTD 46896–97), two males ( MHNSM 24509, MTD 46895); 12 juveniles ( MHNSM 24518, 24523–30, MTD 46898–200), all collected by J. Icochea; one juvenile ( RMNH 27069) collected at the type locality between 29 June and 1 July 1987 by S. B. Hedges.
Referred specimens: two (USNMFS 171096: adult male, 171095: adult female) collected at the type locality between 29 June and 1 July 1987 by S. B. Hedges.
Diagnosis. A medium-sized species of Rhinella attaining a maximum SVL of 45.7 mm in adult females (MHNSM 24510). The new species is distinguished from all congeners by the following combination of characters: (1) canthus rostralis distinct, orbitotympanic and postorbital crests weak ( Fig. 2 View FIGURE 2 ); prominent vertical keel on snout (2) tympanum distinct, oval; (3) parotoid glands moderately large (about same size as ED), subtriangular, not contacting eye; (4) numerous small, tubercles (keratin-tipped in males) scattered on dorsal surfaces of body; (5) forearm moderately long, slim in females, hypertrophied in males ( Fig. 3 View FIGURE 3 ); (6) weakly defined dorsolateral row of slightly enlarged, conical tubercles; (7) tarsal fold absent; (8) webbing on hands and feet fleshy ( Fig. 4 View FIGURE 4 ); (9) first finger shorter than second; (10) males with vocal slits and small keratinous spines on Fingers I and II.
We assign Rhinella yanachaga tentatively to the veraguensis Group sensu Duellman and Schulte (1992) based on its distribution and morphological similarities to species in that group. Two species of the veraguensis Group are known from cloud forests of central Peru ( Fig. 5 View FIGURE 5 )— Rhinella chavin and R. multiverrucosa .
Rhinella yanachaga is easily distinguished from both by being smaller (maximum SVL 45.7 mm vs. 64.9 mm in R. chavin , 68.9 mm in R. multiverrucosa ), by having long, relatively slender extremities with enlarged, conical tubercles (short, stout, with large, flat, elongate warts). Males of R. chavin , R. multiverrucosa , and R. yanachaga have keratinous nuptial pads on Fingers I and II, but R. yanachaga has vocal slits (absent in R. chavin , and R. multiverrucosa ) and hypertrophied arms (not hypertrophied in R. chavin , and R. multiverrucosa ). Other arboreal species of Rhinella include R. arborescandens , R. manu , and R. tacana . Rhinella yanachaga differs from these species by having, a tympanic membrane (absent in R. arborescandens ), and well developed webbing on hands and feet (absent in R. arborescandens ). Rhinella yanachaga differs from R. manu by lacking a tarsal fold (present in R. manu ), by having parotoid glands about as large as eye diameter (larger), a tan venter with brown flecks (red with dark blotches), males with vocal slits (absent), and keratinious
Rhinella yanachaga is most similar to R. tacana from northern Bolivia: both have long, slender extremities, snout with a vertical keel, and males with vocal slits, but R. yanachaga is larger (maximum SVL 45.7 mm vs. 34.2 mm in R. tacana ), has weakly defined dorsolateral rows of slightly enlarged, conical tubercles (dorsolateral row more prominent in R. tacana ), enlarged tubercles on dorsal surfaces of tibia and tarsus, and outer ventrolateral surface of tarsus (extremities without enlarged tubercles), has webbing between Fingers I and II well developed (basal, almost absent), males with keratinous nuptial pads on Fingers I and II (restricted to Finger I), males with hypertrophied arms (arms not hypertrophied), iris olive brown or bronze with black reticulations (iris green).
Description of the holotype. Body moderate to slender; head slightly broader than long; snout slightly pointed in dorsal profile, protruding in lateral profile ( Fig. 2 View FIGURE 2 ); snout with a distinct vertical keel, bearing two small, keratin-tipped tubercles; canthus rostralis more distinct anteriorly than posteriorly, covered with small tubercles each with single keratinized tip, conical in profile; orbitotympanic and postorbital crests distinct, covered with small keratin-tipped tubercles; dorsum of head flat, skin co-ossified with underlying cranial bones; interorbital distance greater than eyelid width; internarial area concave; nostrils protuberant, directed laterally; loreal region concave; lips rounded; small V-shaped notch at symphysis of upper jaw; oral ricti positioned at level of tympanum; tympanum diameter smaller than eye diameter; tympanic annulus a narrow rim anterolaterally, slightly covered by small tubercles on left side of head, dorsal and posteroventral margins indistinct, not in contact with parotoids or postorbital crests; skin on dorsum with numerous small, round, elevated tubercles, conical in profile, bearing single keratinized tip on anterior half of body; posterior half of dorsum less spinous and with larger tubercles than on anterior half of body; parotoid glands ovoid in dorsal view, subtriangular in dorsolateral view, longer than wide, widest posteriorly, slightly smaller than upper eyelid, descending onto side of head without contacting eye; upper eyelid with conical keratin-tipped tubercles, largest on outer margin.
Flanks with lower density of tubercles than dorsum, but with few, slightly enlarged tubercles (left side five tubercles, right side two tubercles) between arm and leg insertion, not forming a distinct dorsolateral fold; skin of throat, chest, and venter granular; granules on venter larger than those on chest and throat; ventrolateral row of enlarged tubercles absent; arms long, hypertrophied ( Fig. 3 View FIGURE 3 A); hands relatively large with long fingers; relative length of adpressed fingers 1 <2 <4 <3; webbing fleshy; webbing formula of fingers I1–2II2– 3III3–3 IV ( Fig. 4 View FIGURE 4 A); lateral fringes broad; tips of digits terminating in indistinct discs; subarticular tubercles ovoid in ventral aspect, flat in profile; supernumerary tubercles numerous, about 25% as large as subarticular tubercles; palmar tubercle large and round, flat in profile, about four times size of thenar tubercle; thenar tubercle ovoid, round in profile; limbs long, slim; tibia length shorter than foot length; tarsal fold absent; inner metatarsal tubercle ovoid, flat in profile, twice and a half the size of outer metatarsal tubercle; outer metatarsal tubercle ovoid, conical in profile; toes relatively long; relative length of adpressed toes 1 <2 <3 <5 <4; fleshy webbing; webbing formula of toes I1–1 II1–11 /2 III1–21 /2 IV3–2 -V ( Fig. 4 View FIGURE 4 B); broad lateral fringes present; subarticular tubercles ovoid in ventral aspect, flat in profile; supernumerary tubercles numerous about 35% as large as subarticular tubercles.
Skin texture of dorsal surface of forearms as on dorsum; dorsal surface of tibia with enlarged tubercles (4– 6 times the size of tubercles on dorsum), ovoid in dorsal view, conical in profile; ventral surface and outer ventrolateral surface of tarsus with many enlarged round to ovoid tubercles, conical in profile; outer dorsolateral margin of foot with numerous enlarged round to ovoid tubercles; cloacal opening protuberant, directed ventrally near lower level of thighs ( Fig. 3 View FIGURE 3 C); inguinal fat bodies absent; choanae small, ovoid; maxillary, premaxillary, mandibular, and vomerine teeth absent; tongue elongate, three times as long as wide, about equal in width throughout its length, free posteriorly for about two fifths its length; vocal slits nearly straight, located bilaterally at posterior half of mouth floor between tongue and margin of jaw.
Measurements (in mm) of holotype: SVL: 38.4; TL: 12.8; FL: 16.4; HL: 11.3; HW: 12.0; ED: 2.1; TY: 1.2; IOD: 3.6; EW: 2.5; IND: 2.5; E–N: 2.8; PL: 4.0; PW: 2.9.
Coloration of holotype in preservative: Dorsum brown, parotoids brownish orange; narrow tan middorsal stripe extending from snout to cloaca; broad, tan dorsolateral stripe on each side of flanks extending from parotoids to groin; flanks ventrolaterally with weakly defined broad, brown stripe beginning behind tympanum and extending to inguinal region; tympanum dark brown; upper lip cream with dark brown bar below eye, and below tympanum; throat, chest and venter tan with minute grayish-brown spots; ventral surfaces of hands and feet gray. Coloration of holotype in life unknown.
Variation. Descriptions of coloration in life for an adult male, adult female, and a juvenile are based on field notes by S. B. Hedges: The adult male (USNMFS 171096, Fig. 6 View FIGURE 6 ) was very dark brown dorsally with small, irregular, green spots and markings; most situated just above the dorsolateral row of tubercles and with one large green blotch on the right side just behind the parotoid glands. The sides (below dorsolateral row of tubercles) were reddish brown and the upper lip was reddish brown with green flecks. The ventral surface was orange brown and had small green spots on the chin, darker brown flecks on the belly, and very dark brown or black markings under the limbs. The iris was olive brown. The female (USNMFS 171095) was tan dorsally with light brown markings. The narrow middorsal stripe was yellowish with small irregular black markings on each side. The sides were dark brown with a greenish spot posterior to the tympanum and above the axillary region. The ventral surface was mottled with brown (reddish brown on chest) and had a faint yellowish midventral line. The eyes were bronze.
The type series shows some variation in coloration pattern: Four (MHNSM 24520, 24522, 24526, USN- MFS 171095) specimens have a tan middorsal stripe as described for the holotype, and several specimens (e.g., MHNSM 24511, 24514, MTD 46899-900) have small, ovoid black flecks on the dorsum, and others (e.g., MHNSM 24511, 24518, 24525, MTD 46899) a dark brown X-shaped blotch on head and shoulder. Ventral coloration varies from tan with few grayish brown flecks (MHNSM 24517) to tan and brown mottled (MTD 46895) to dark brown with tan flecks (MHNSM 24510).
There is obvious sexual dimorphism in skin texture: Dorsal skin of males is spinose because of keratintipped tubercles; skin of females smooth without keratin-tipped tubercles. Furthermore, all males have vocal slits, nuptial pads on dorsal and inner lateral surfaces of thumb and on dorsal surface of second finger, and arms are hypertrophied, whereas arms of females are slim ( Fig. 3 View FIGURE 3 A, B). The cloaca is more protuberant in males than in females and the opening is directed ventrally in males, whereas in females laterally ( Fig. 3 View FIGURE 3 C, D). Females are slightly larger than males (maximum SVL in females 45.7 mm vs. 41.6 mm in males).
Crests are more prominent in adult than in juvenile specimens, and are most prominent in an adult female (MHNSM 24513). All have the snout with a distinct vertical keel which is more prominent in adults. The dorsolateral row of tubercles is generally weakly defined or indistinct, as the tubercles do not differ much in size compared to other dorsal tubercles, but in some specimens (MHNSM 24510) is the dorsolateral row is more distinct as the tubercles coalesce. For measurements of the type series of adult R. yanachaga , see Table 1 View TABLE 1 ; for ranges and proportions see Table 2 View TABLE 2 .
Comparative cranial osteology. The skull of Rhinella yanachaga generally resembles those of other R. veraguensis Group species, but certain cranial features of this taxon are notably different. Detailed comparative osteological descriptions and illustrations for other species in the R. veraguensis Group are provided elsewhere ( Lehr et al. 2005, Pramuk 2006, Chaparro et al. 2007). The skull of R. yanachaga is broadly rounded anteriorly and is wider than long with the greatest width being at the level of the quadratojugals ( Fig. 7 View FIGURE 7 A–C). The dorsal surfaces of the dermal bones are relatively smooth and are not exostosed or ornamented with pits and rugosities (as they are in R. multiverrucosa and to a lesser extent in R. chavin and R. veraguensis ). Of the five veraguensis Group species examined osteologically (Appendix 1), the skull of R. yanachaga is most similar in structure to the crania of R. chavin , R. manu , and R. veraguensis and in overall appearance and skull proportions, this species most closely resembles R. manu . Both of these species have similarly broad, rectangular, and flattened frontoparietals broadly contacting the medial surfaces of the otic rami of the squamosals (forming a complete temporal arcade sensu Lynch 1971). In contrast, the frontoparietals of R. veraguensis do not contact broadly the otic rami and are slightly expanded dorsolaterally, creating relatively well-developed supraorbital crests. In R. yanachaga , supraorbital crests are weakly developed. Relative to most other bufonids, species of the R. veraguensis Group have relatively lightly to moderately ossified skulls, a consequence of which is exposure of the dorsal surface of the sphenethmoid. The sphenethmoid of R. yanachaga is exposed to a greater extent than it is in other species of this group. The nasals and frontoparietals barely contact one another laterally whereas they do in all other species examined, yielding a distinctively triangular appearance to the exposed surface of the sphenethmoid. In R. yanachaga , R. manu and R. veraguensis , only the lateral-most edges of the premaxillae are visible in dorsal view (in R. chavin , and R. multiverrucosus , the In ventral view ( Fig. 7 View FIGURE 7 B), the premaxillae and maxillae of R. chavin , R. multiverrucosa , and R. yanachaga are deeper and more robust than those of R. veraguensis . Moreover, in dorsal view the maxillae of R. chavin , R. manu , R. multiverrucosa , and R. yanachaga are angled posterolaterally to a greater degree from their point of contact with the premaxillae than they are in R. veraguensis yielding a relatively more rounded appearance to the maxillary arcade of this species. Rhinella yanachaga , R. chavin R. manu , and R. multiverrucosa have a narrow ridge present on the ventral surface of the parasphenoid that extends from the point of contact of the medial ramus of the pterygoid and parasphenoid to the medial point of the parasphenoid. A ridge on the parasphenoid also is present in R. veraguensis ; however, it is only present medially and does not extend to the point where the pterygoid contacts the parasphenoid. In ventral view, the anterior edge of the cultriform process of the parasphenoid is dramatically truncated (similar to the condition seen in R. manu ); in other species it is relatively more acuminate with R. veraguensis displaying the most acuminate condition. The vomers are robust in R. veraguensis and have distinctive postchoanal and prechoanal processes; whereas, in R. yanachaga , R. chavin R. manu , and R. multiverrucosa , the vomers are more slender and lack well-developed postchoanal processes.
In lateral view ( Fig. 7 View FIGURE 7 C), R. yanachaga has a relatively shallow braincase similar to that of R. veraguensis (compared to the relatively deep skulls of R. chavin and R. multiverrucosa ). As with all members of the R. veraguensis Group, the posterior part of the braincase, including the occipital condyles, extends well beyond the posterior edge of the jaw symphysis; columellae are present and columnar. The quadratojugals of R. yanachaga are relatively small like those of R. veraguensis and R. manu (compared to the relatively robust quadratojugals of R. chavin and R. multiverrucosa ). Unlike other members of the veraguensis Group, the anteroventral edge of the nasal of R. yanachaga abuts the anterior edge of the maxilla.
Distribution and ecology. Rhinella yanachaga is only known from the Cordillera Yanachaga in the National Park Yanachaga Chemillén ( Fig. 5 View FIGURE 5 ). This park contains the Cordillera de Yanachaga that reaches an elevation of 3643 m, and descends into the valley of the Río Palcazu. The park extends westward to the Cordillera de Santa Bárbara reaching an elevation of 3400 m and is separated from the Cordillera de Yanachaga by the deep canyon of the Río Huancabamba. Thus, Rhinella yanachaga occurs in a protected region that encompasses forested slopes of the Andes. The type locality is reached from Oxapampa by trail, as described in Hedges (1990). Habitat at the type locality is cloud forest with a thick layer of moss covering the ground and tree trunks. Three specimens (RMNH 27069, USNMFS 171095, 1710096), were collected between 29 June and 1 July 1987 at night. One adult male (USNMFS 171096) was found beneath a layer of moss and dirt about 10–20 cm below the surface on the side of the trail. The other (USNMFS 171095) was found resting on the tops of leaves (ca. 50 cm above the ground) along the trail. Other specimens were found in trees (J. Icochea, personal communication). Syntopic species include Phrynopus bracki , Pristimantis mendax , and P. sagittulus .
One gravid female (MHNSM 24510, SVL 45.7 mm) was dissected and contained 136 (left ovary: 66; right ovary: 70) unpigmented eggs, which are pale orange in preservative, similar to those of Pristimantis or Phrynopus . The call and tadpole of R. yanachaga are unknown.
Etymology. The specific name yanachaga is derived from Quetchuan and means black. The name is used as a noun in apposition and refers to the Cordillera Yanachaga where the new species was found.


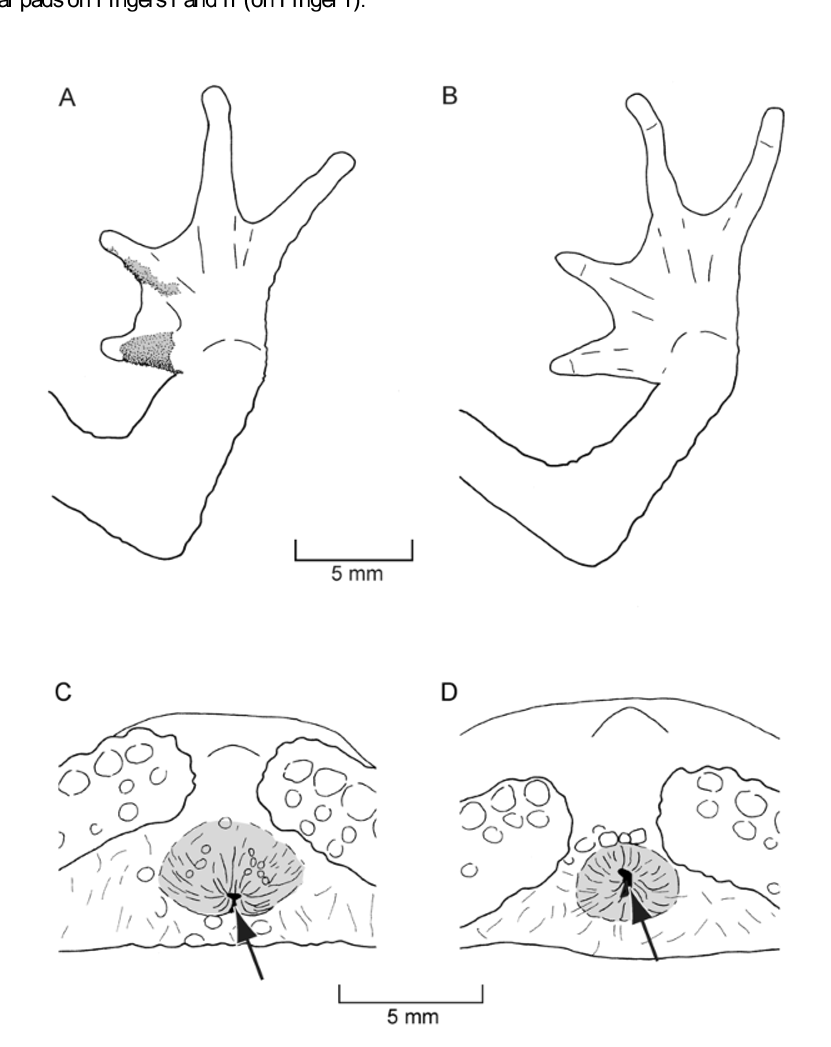



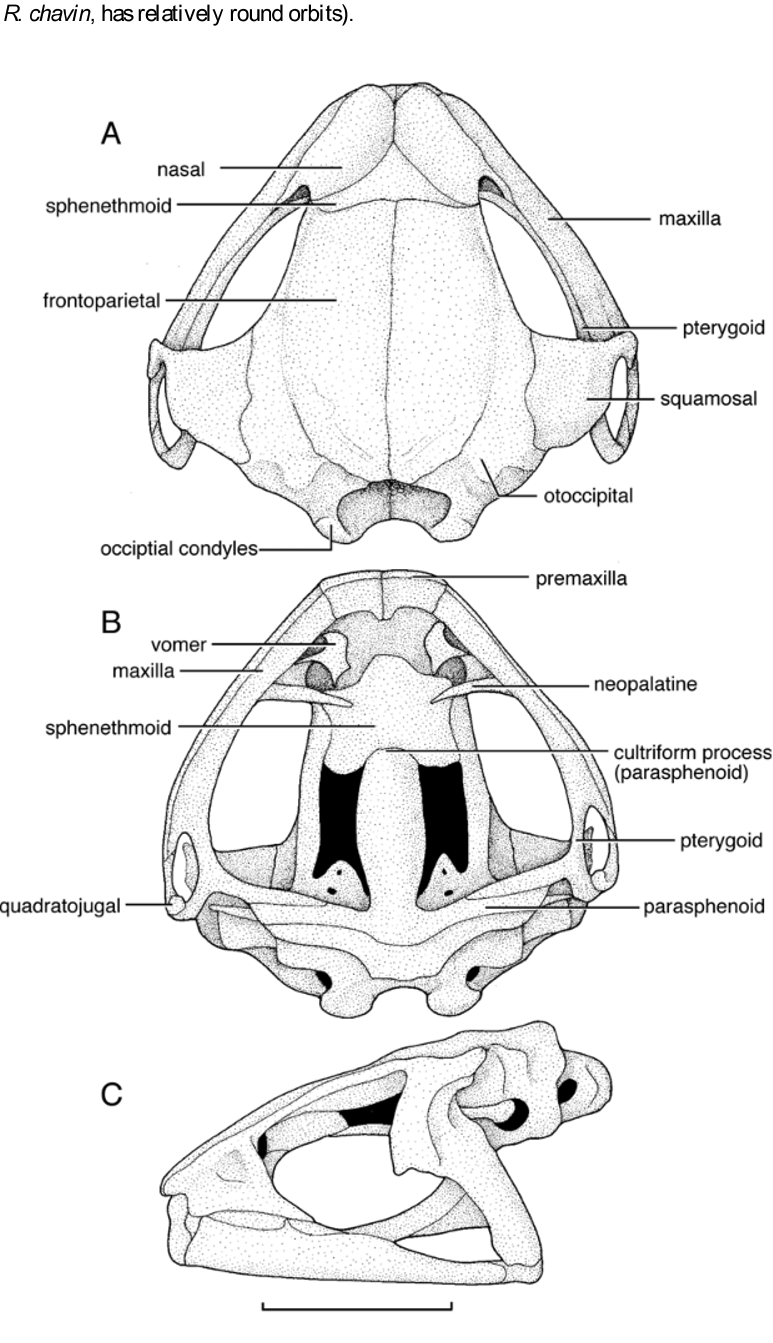
TABLE 1. Measurements (in mm) of selected adult paratypes of Rhinella yanachaga.
| character | MHNSM 24510 | MHNSM 24509 | MTD 46895 | MHNSM 24511 | MTD 46896 | MHNSM 24512 | MHNSM 24513 | MHNSM 24515 |
|---|---|---|---|---|---|---|---|---|
| Sex | f | m | m | f | f | f | f | f |
| SVL | 45.7 | 41.6 | 41.0 | 37.9 | 36.9 | 36.5 | 36.4 | 35.8 |
| TL FL | 15.7 19.4 | 14.3 18.6 | 14.5 18.3 | 13.4 17.6 | 12.4 15.9 | 13.4 16.4 | 13.0 17.2 | 12.0 15.7 |
| HL | 14.6 | 13.4 | 13.7 | 12.9 | 12.1 | 12.6 | 13.9 | 11.9 |
| HW | 13.9 | 13.4 | 12.6 | 12.3 | 11.4 | 12.3 | 12.4 | 11.1 |
| ED | 3.2 | 2.6 | 2.3 | 3.1 | 2.6 | 2.8 | 3.2 | 2.6 |
| TY | 2.6 | 2.1 | 1.9 | 1.7 | 1.8 | 2.2 | 2.0 | 1.7 |
| IOD EW | 4.5 3.2 | 3.9 3.6 | 3.8 3.7 | 4.5 3.0 | 3.9 3.2 | 4.3 3.5 | 4.0 3.0 | 4.0 3.0 |
| IND | 3.3 | 2.9 | 3.0 | 2.9 | 2.6 | 3.2 | 2.9 | 2.8 |
| E–N | 3.6 | 3.8 | 3.5 | 3.9 | 3.5 | 3.5 | 3.6 | 3.2 |
| PL | 3.5 | 5.0 | 5.4 | 3.6 | 3.7 | 3.8 | 3.3 | 4.0 |
| PW | 4.0 | 3.9 | 4.2 | 3.5 | 3.9 | 4.3 | 3.8 | 3.5 |
TABLE 2. Ranges (in mm) followed by mean and standard deviation and proportions of adults of Rhinella yanachaga.
| Characters | Females (n = 14) | Males (n = 3) |
|---|---|---|
| SVL | 29.1–45.7 (35.6±3.7) | 38.4–41.6 (40.3± 1.7) |
| TL | 10.0–15.7 (12.6±1.3) | 12.8–14.5 (13.9±0.9) |
| FL | 12.0–19.4 (15.9 ±1.9) | 16.4–18.6 (17.8±1.2) |
| HL | 10.1–14.6 (12.3 ±1.2) | 11.3–13.7 (12.8±1.3) |
| HW | 9.5–13.9 (11.7±1.1) | 12.0–13.4 (12.7±0.7) |
| ED | 2.1–3.2 (2.8± 0.3) | 2.1–2.6 (2.3± 0.3) |
| TY | 1.0–2.6 (1.6±0.4) | 1.2–2.1 (1.7±0.5) |
| IOD | 3.4–4.5 (4.0 ±0.4) | 3.6–3.9 (3.8±0.2) |
| EW | 2.3–3.5 (3.0±0.4) | 2.5–3.7 (3.3± 0.7) |
| IND | 2.2–3.3 (2.8± 0.3) | 2.5–3.0 (2.8±0.3) |
| E–N | 2.8–4.0 (3.4± 0.4) | 2.8–3.8 (3.4± 0.5) |
| PL | 2.9–4.3 (3.5±0.5) | 4.0–5.4 (4.8±0.7) |
| PW | 2.9–4.3 (3.6± 0.4) | 2.9–4.2 (3.7± 0.7) |
| TL/SVL | 0.33–0.38 | 0.33–0.35 |
| FL/SVL | 0.40–0.51 | 0.43–0.45 |
| HL/SVL | 0.29–0.38 | 0.29–0.33 |
| HW/SVL | 0.30–0.37 | 0.31–0.32 |
| HW/HL | 0.89–1.06 | 0.92–1.06 |
| E–N/ED | 1.10–1.44 | 1.33–1.52 |
| EW/IOD | 0.68–0.82 | 0.69–0.97 |
| TY/ED | 0.41–0.81 | 0.57–0.81 |
| PL/ED | 0.93–1.90 | 1.90–2.35 |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.