
Triops cf. granarius ( Lucas, 1864 )
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4446.4.8 |
|
publication LSID |
lsid:zoobank.org:pub:B55AD7EB-CD21-4AB5-9BBA-1C85449A8B73 |
|
DOI |
https://doi.org/10.5281/zenodo.5955718 |
|
persistent identifier |
https://treatment.plazi.org/id/039987B6-FF86-FFD4-A0DA-F9F0FE5C9617 |
|
treatment provided by |
Plazi |
|
scientific name |
Triops cf. granarius ( Lucas, 1864 ) |
| status |
|
Triops cf. granarius ( Lucas, 1864) View in CoL from Socotra
( Figs 2–5 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 )
Material examined. Three specimens were examined for this study, two adult females and one adult male, reared from incubated sediment that was collected 28.I.2014 at Eriosh (alt. 20m, coordinates N12° 35.497’ E53°49.048’ or N12.5916167° E53.817467°), Socotra Island ( Yemen) by KVD (pers. coll.).
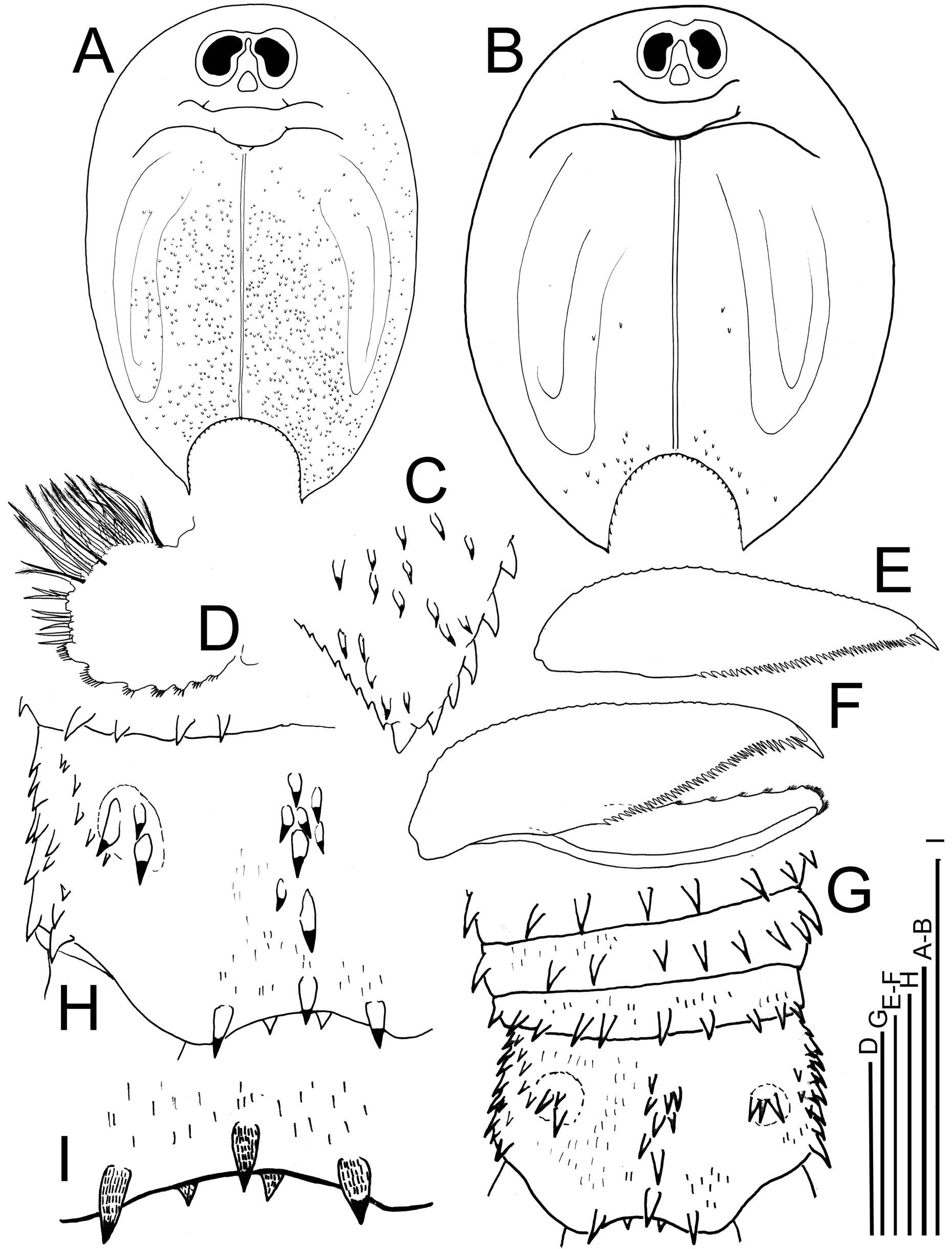
Characters of the T. granarius species group. The absence of a supra-anal plate, visible with the naked eye ( Fig. 2B View FIGURE 2 ), places these notostracans in Triops . Morphological characters that are considered diagnostic for the T. granarius species complex in the classical sense (sensu Longhurst 1955a) are present in the Socotran specimens. These features include: i) dorsal (nuchal) organ triangular, its anterior margin reaching between the compound eyes, not situated posterior to the eyes ( Figs 2B View FIGURE 2 & 5A–B View FIGURE 5 ); ii) carina with none (or one undeveloped) spine anterior to the terminal spine ( Figs 3C–D View FIGURE 3 & 4D–E View FIGURE 4 ); iii) supernumerary spines present on the ventral face of the apodous segments ( Fig. 3E View FIGURE 3 ); iv) telson dorsal margin armature with various median spines in an irregular row, which are not large ( Figs 5G–H View FIGURE 5 ); telson armature variation in the T. granarius species complex is shown in Hamer & Rayner (1995) and Longhurst (1955a); v) number of apodous segments (two females): 11–13, male 13; vi) second maxilla present ( Fig. 5D View FIGURE 5 ). Females were distinguished from the male, using the mystax and the P11 (modified in females).
Morphological notes on the Socotran population. Length (without furca): 29,1mm (male), 23 and 27,2mm (females); Number of body segments (not including telson): 43 of which 13 apodous (male); 41 (11 apodous) and 43 (13 apodous) in the two female specimens. Colour brown to greenish ( Figs 2B View FIGURE 2 & 3 View FIGURE 3 ).
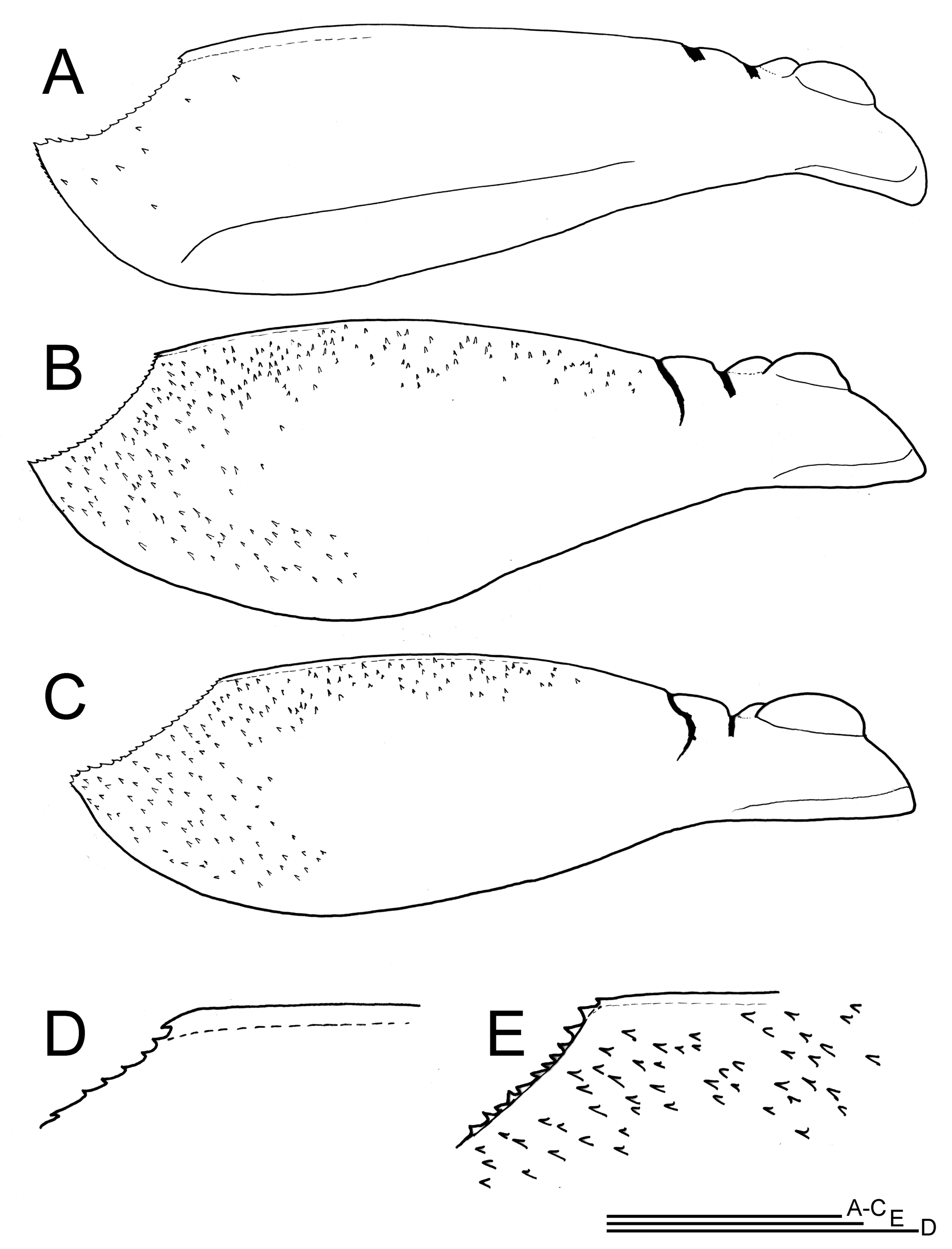
Carapace (lateral view in figs 3A–B & 4A–C; dorsal view in figs 2B & 5A–B) length/width (length from anterior margin to posterior apices of carapace sulcus): 19/ 15mm (male), 17,5/ 13mm and 19/15,1mm (females). In dorsal view, the carapace is wider in posterior part in the male than in females, more tapering in the females and giving the male a more round appearance ( Figs 5A–B View FIGURE 5 ; however only three specimens to compare). In lateral view, the carapace length is about 3–3.5 x height (at highest point) ( Figs 3A–B View FIGURE 3 & 4A–C View FIGURE 4 ). Female carapace posterior surface covered in minute spines ( Figs 3B View FIGURE 3 , 4B–C View FIGURE 4 & 5A View FIGURE 5 ), males with only a few scattered spines ( Figs 3A View FIGURE 3 , 4A View FIGURE 4 & 5B View FIGURE 5 ). Short spines on the carapace carina suggests the trachyaspis -form, a known morphological variation in T. granarius ( Longhurst 1955a) and apparently it is variable here in one population hatched from sediment. The single male studied here lacked this feature, except for a few scattered spines on the posterior surface ( Fig. 5B View FIGURE 5 ), whereas the two females showed a carapace covered in spines ( Figs 2B View FIGURE 2 , 3B View FIGURE 3 , 4B–C View FIGURE 4 , 5A&C View FIGURE 5 ). These spines have a chitinized tip ( Fig. 5C View FIGURE 5 ). Carapace sulcus ( Figs 2B View FIGURE 2 & 5A–B View FIGURE 5 ) spines ca. 40–50 (females), 40 (male). Sulcus in male about as deep as wide (4mm), in females about two times as deep as wide (2–3mm wide, 4–5mm deep) ( Figs 4A– C View FIGURE 4 ). Carina on carapace well developed, terminal spines with one small or no anterior teeth, further no carinal spines ( Figs 3C–D View FIGURE 3 & 4D–E View FIGURE 4 ). Male carina more robust with the posterior end directed ventrally, whereas in females it remains straight ( Figs 3D View FIGURE 3 versus 3C & 4B–C versus 4A). In general, the entire dorsum is slightly convex ( Figs 3A–B View FIGURE 3 & 4A–C View FIGURE 4 ).
Eyes (dorsal view Figs 2B View FIGURE 2 & 5A–B View FIGURE 5 , lateral view Figs 3A–B View FIGURE 3 & 4A–C View FIGURE 4 ) well developed, strongly protruding in lateral view; eye field with strong constriction in middle ( Figs 5A–B View FIGURE 5 ). Dorsal organ triangular with rounded corners, its anterior margin reaching just beyond the posterior margins of the eyes ( Fig. 5A–B View FIGURE 5 ).
Labrum ( Fig. 3F View FIGURE 3 ) rectangular.
Second maxilla ( Fig. 5D View FIGURE 5 ) well developed and round, longest setae on the margin less than the diameter of the structure. Row of spiniform setae on the margin continuous, joined by anterior slender setae in the second half of the structure.
Thoracopods (only some parts studied) P2 endopodite ( Fig. 5E View FIGURE 5 ) elongate, about 3.5 times as long as wide and tapering distally and well developed distal claw, straight and about twice the length of the adjacent anterior spine; posterior margin of endopodite implanted with spines just over half of its length, these spines are not equal in size (irregular, some are a third to half shorter). P6 endopodite ( Fig. 5F View FIGURE 5 ) elongate and tapering, about 3x as long as wide and with a pointed apex; distal claw large, curved inwards, and with strongly chitinized tip, twice the length of adjacent spine; spines implanted to about half the posterior margin, four to five distalmost spines, close to distal claw, largest.
Apodous segments with supernumerary spines present on ventral side, clear in the male, about 8–13 per segment ( Fig. 3E View FIGURE 3 ); in females there are fewer supernumerary spines seen per segment (4–8).
Telson armature ( Figs 5G–I View FIGURE 5 ) median spines eight and small, forming an irregular row; anterior lateral spines in groups of two to four; posterior marginals relatively small, about 2,5–3 times as long as wide ( Figs 5H–I View FIGURE 5 ) and implanted with small hairs; additional secondary posterior marginal present ( Fig. 5I View FIGURE 5 ). Lateral margins of telson implanted with conical spines, and two posterior furcal spines, about as long as wide ( Fig. 5H View FIGURE 5 ).
Remarks. At this point, it is best to assign the Socotran populations to Triops granarius sensu lato, in the clear understanding that this is a species group which needs revision and that the animals are not T. granarius sensu stricto. The use of the name rather than assigning these animals to one of many available synonyms described from the adjacent land or to a new name is temporary until a more detailed investigation. It is possible that this is a new species. In particular, molecular analysis and examination of more specimens will help to explore the relationship of the Socotran animals. The type locality of T. granarius s. str. is Beijing ( China), the East Asian populations likely forming a separate clade (Korn & Hundsdoerfer 2006, 2016), therefore there is a high chance that the Socotran populations may be assigned to another name in the future. However the assignment is beyond the scope of this record and would best include other Arabian and African T. granarius -like populations in comparison.
From the few limb features examined here, it seems that the Socotran specimens have a closer affinity to the African, not to the Asian clade. The endopodites of the limbs in the Asian populations are suggested to have a more rounded appearance ( Longhurst 1955a), and the African more elongate and acute ( Korn & Hundsdoerfer 2016). In the specimens from Socotra, this structure is clearly elongate and sharp ( Figs 5E & F View FIGURE 5 ). However this character has not been illustrated in literature for the Asian populations and has not been part of any larger formal systematic revisions so far.
A brief examination of the endopodite shape in “true” Asian T. granarius based on photos of trunk limbs (P3– 8) kindly provided by Dr M. Korn (unpubl. data), shows that this is a highly recognisable feature, unmistakably different from the elongate sharp endopodites in the Socotran animals. Furthermore it is clear that the Socotran animals are distinctly different from the Moroccan species that were split by Korn & Hundsdoerfer (2016) from the T. granarius group, in the setulation of the second maxilla (much shorter in Socotran populations), the endopodite spines (more irregular in the Moroccan species) and the telson armature (secondary posterior marginal spines present in Socotran animals). Unfortunately other members of the T. granarius group have not been described or redescribed adequately and the morphological notes on the populations from the Arabian mainland shown in Thiéry (1996) do not provide enough details for comparison. The only feature Thiéry (1996) mentions are the number of apodous body segments in the Omani female T. granarius specimens as being less (8–9) than the Saudi populations (9–12). On Socotra the females have more (11–13) apodous body segments but this feature is variable and only two female specimens were examined so far.
Local ecology. Triops is found with two other large branchiopods that hatched from the sediment, the endemic anostracan Branchipodopsis relictus and a yet unidentified spinicaudatan. Among the small branchiopods, the endemic Moina diksami Van Damme & Dumont, 2008 and an unidentified Daphnia ( Ctenodaphnia ) sp. were observed (ephippia). No details on the ecological characteristics of the site were collected, except that a series of connected large shallow temporary pools or a very shallow temporary lake forms after rains on the limestone. Dry sediment was stored in the dark at room temperature for two years and Triops hatched quickly under constant light conditions, appearing 24–48 hrs after wetting the sediment with bottled spring water at constant room temperature. Branchipodopsis hatched simultaneously and raced Triops to adulthood; notostracans reached maturity after about two weeks, the anostracans after four to six days.
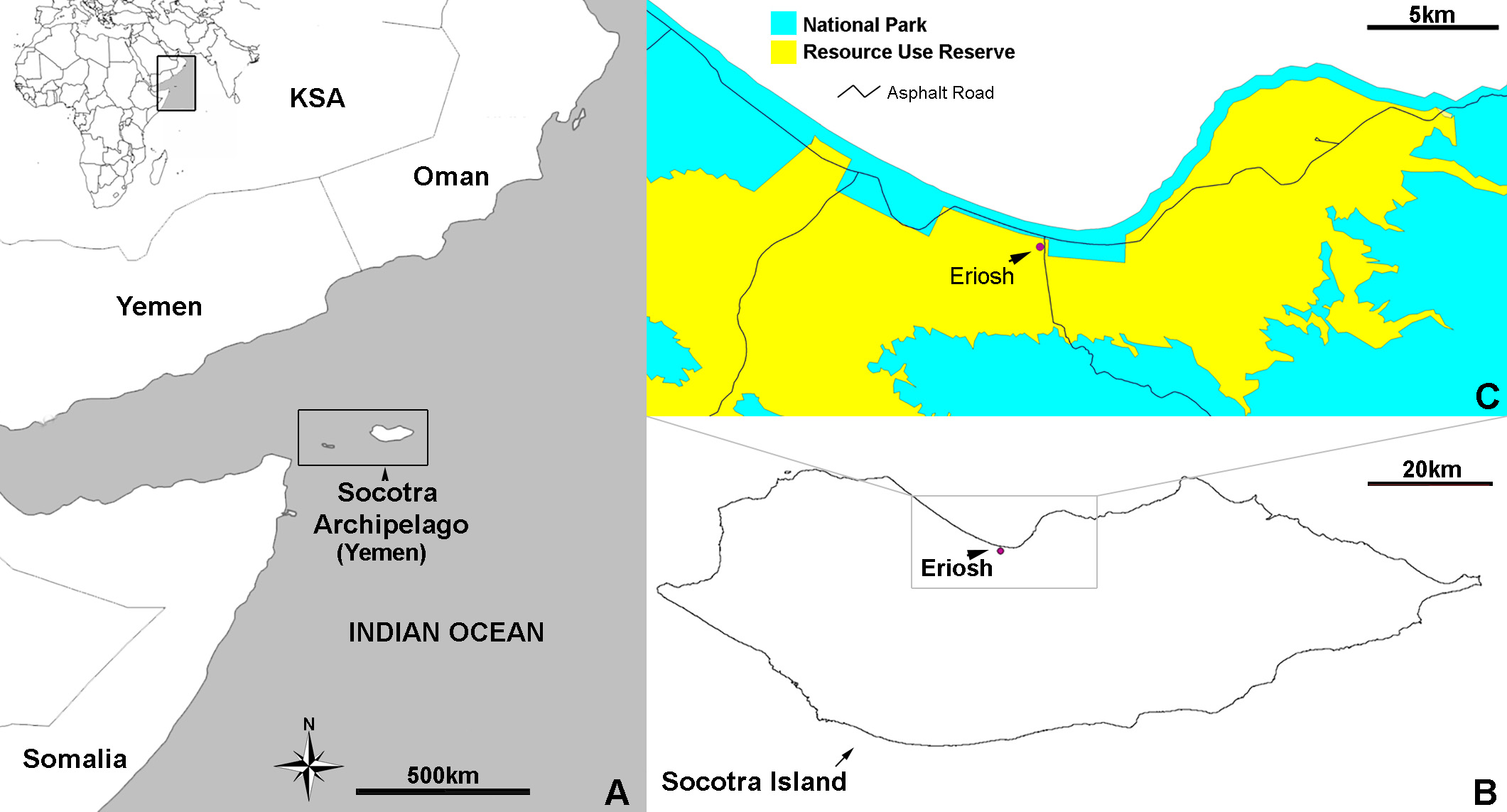
Distribution/Biogeography. Local. So far only one locality on Socotra Island is known to harbour Triops . Considering that Eriosh has been used traditionally as a watering place for camels and goats during rainy seasons ( Doe, 1970) and taking the strong winds on Socotra into account ( Cheung & DeVantier 2006, Scholte & De Geest 2010), it is likely that Triops may be found elsewhere on the island. Suitable habitats would be available where larger temporary pools are formed.
Global. T. granarius s. str. was originally described from Beijing, China. The species group has the largest geographical and morphological range among the four major lineages in the genus ( Longhurst 1955a, Korn & Hundsdoerfer 2006, 2016). Considering the growing evidence for high diversity in this species group, the Socotran animals may well belong to a different species. The complex is widespread in the Old World and the animals are found in suitable habitats from Japan to South Africa ( Longhurst 1955a). Triops is well represented in several islands around the world ( Longhurst 1955b), Mediterranean ( Lanfranco et al. 1991; Marrone et al. 2006, Tzirotzis et al. 2014), Caribbean and the Galápagos ( Linder 1960) and Madagascar ( Longhurst 1955a). The high dispersal capacity of the dormant eggs allows Triops to occupy temporary pools in remote islands such as Hawai’i and New Caledonia ( Longhurst 1955a – b). The Socotran population is the first evidence of T. granarius (and only the second of Triops ) from islands in the Indian Ocean.
Importance of conservation of the Triops locality. There is no formal protection in place in Socotra for freshwater ecosystems. The locality falls in a rapidly conserved coastal area and in a zone with moderate protection as designed in the original Socotra Conservation Zoning Plan (Resource Use Reserve; EPC/UNDP GEF, 2000). Eriosh is not part of the Socotra National Park, yet just adjacent (<1km; Fig. 1C View FIGURE 1 ) to it, and therefore falls on the border of UNESCO terrestrial core and buffer properties. The site is located next to the junction of two major roads (also <1km) on the island and has been damaged by the construction of the Diksam road, which cuts through the centre of the area, destroying a considerable part of it ( Van Damme et al. 2004, Van Damme & De Geest 2006). However, the Socotra Resource Use Reserve legally aims to “ protect and maintain the unique biological diversity of Socotra islands, as well as other natural, cultural and landscape values of the area in the long term ” and in general, the Zoning Plan aims to “ protect the genetic material of rare and endemic species in Socotra islands ” (Articles 2.5 and 4a in the Zoning Plan; EPC/UNDP GEF 2000). It is therefore important here to briefly highlight natural values and the existence of rare populations (or species) such as an insular Triops in the western Indian Ocean, for conservation purposes and to mention the importance of a neglected habitat such as temporary pools.
From a biological diversity perspective, as this is the only place on the island know so far where three large branchiopods are found together and the only locality known in an Indian Ocean island besides Madagascar that contains Triops , this locality can be considered as important. Furthermore, considering the large cryptic diversity in the T. granarius species group, there is a possibility that the Socotran Triops may belong to a new species. The anostracan Branchipodopsis in this locality is an endemic, so is the cladoceran Moina Baird, 1850 , while other interesting taxa (Spinicaudata and Daphnia) are yet unidentified. The habitat should be locally protected, if only for the branchiopods.
For additional protection beyond the existing Socotra Conservation Zoning Plan, Eriosh could be designated as a Site of Special Scientific Interest (SSSI) or its local equivalent in future conservation management plans on Socotra, to avoid the risk of losing its value for biodiversity. Immediate threats to this locality include the direct destruction through infrastructure development, which is expanding in Socotra along the coastal roads, and pollution ( Van Damme & Banfield 2011). Without formal protection there is a high probability that this locality, like much of the northern coastal plain, may be affected in the coming decades, as much of the coastal areas on Socotra are being bought up by foreign investors for development regardless of the biodiversity value of this UNESCO World Heritage site. The typical habitats of Triops , ephemeral pools, are rarely incorporated into conservation measures in tropical or subtropical regions in general. However, they play an important role as model systems in conservation biology ( Wiggins et al. 1980, De Meester et al. 2005) and the habitat is increasingly making way to land development globally ( Céréghino et al. 2008, Williams et al. 2001). In Europe the conservation of this neglected habitat and in particular of Triops populations, is taken seriously as these large branchiopods have become endangered and have gone locally extinct in most countries ( Eder & Hödl 2002, Brendonck et al. 2008, Gołdyn et al. 2012). In the UK for example, Triops cancriformis is classified as Endangered under the Wildlife and Countryside Act and the only two localities where it is still found, are under legal protection ( Hughes 1997, Feber et al. 2011, Sellers et al. 2017). There is even a Triops Conservation Group in the UK which also coordinates ex-situ breeding of the population ( Hughes 1997).
The conservation of this particular locality which harbours Triops in Socotra may not only be important from the perspective of biodiversity, evolution and biogeography, but perhaps from a cultural perspective as well. These animals may actually contribute to protecting the unique petroglyphs on the island. Because of their active foraging and burrowing habits and the large numbers in which they occur, tadpole shrimps are able to modify the aquatic environment they live in through bioturbation as they stir up sediments (as ecosystem engineers; Croel & Kneitel 2011). Archaeologists suggest that the main reason for which the surface rock art has not been destroyed naturally throughout the centuries is because of the fine sediment that is distributed over the area each monsoon, effectively forming a small protective layer on the soft limestone against erosion (and looters) as well as the absence of shrubs on the site (J. Jansen van Rensburg pers. comm.). It is possible that Triops , as an active aquatic ecosystem engineer, plays a role in redistributing fine sediment over the inundated rock face after rains and actively uproots seedlings, and may have done so for a very long time. Future development plans for the site should take into account that the dry sediment contains the valuable branchiopod egg bank and should aim to preserve the temporary pools which have an added cultural value as a watering place for herders. Protecting the natural value of the site by conservation of the habitat of the large branchiopods would in this case also protect the cultural values and vice versa. Triops as an ancient guardian of an archaeological treasure.
Future conservation efforts on Socotra may take an innovative role in Arabia by incorporating examples of the invertebrate freshwater faunas and their ecosystems into future management plans, in particular by including widely ignored habitats such as temporary pools which harbour a unique biodiversity. The potential of Triops in the biological control of mosquito larvae, as well as the potential role in local environmental education, may help in awareness and in establishing a locally positive image for these fascinating large branchiopods. Large branchiopods may be locally promoted on Socotra as flagship species in conservation, to help protect their ephemeral habitats. Why not?


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |