
Dipolydora vietnamita, Radashevsky, 2022
|
publication ID |
https://doi.org/ 10.11646/zootaxa.5162.2.1 |
|
publication LSID |
lsid:zoobank.org:pub:11BA9195-9A8E-4098-975B-E010627F9DFD |
|
DOI |
https://doi.org/10.5281/zenodo.6809883 |
|
persistent identifier |
https://treatment.plazi.org/id/EB418B1C-0919-4E41-AAF0-624C42743768 |
|
taxon LSID |
lsid:zoobank.org:act:EB418B1C-0919-4E41-AAF0-624C42743768 |
|
treatment provided by |
Plazi |
|
scientific name |
Dipolydora vietnamita |
| status |
sp. nov. |
Dipolydora vietnamita View in CoL sp. nov.
http://zoobank.org:act: EB418B1C-0919-4E41-AAF0-624C42743768
( Figs 11−14 View FIGURE 11 View FIGURE 12 View FIGURE 13 View FIGURE 14 )
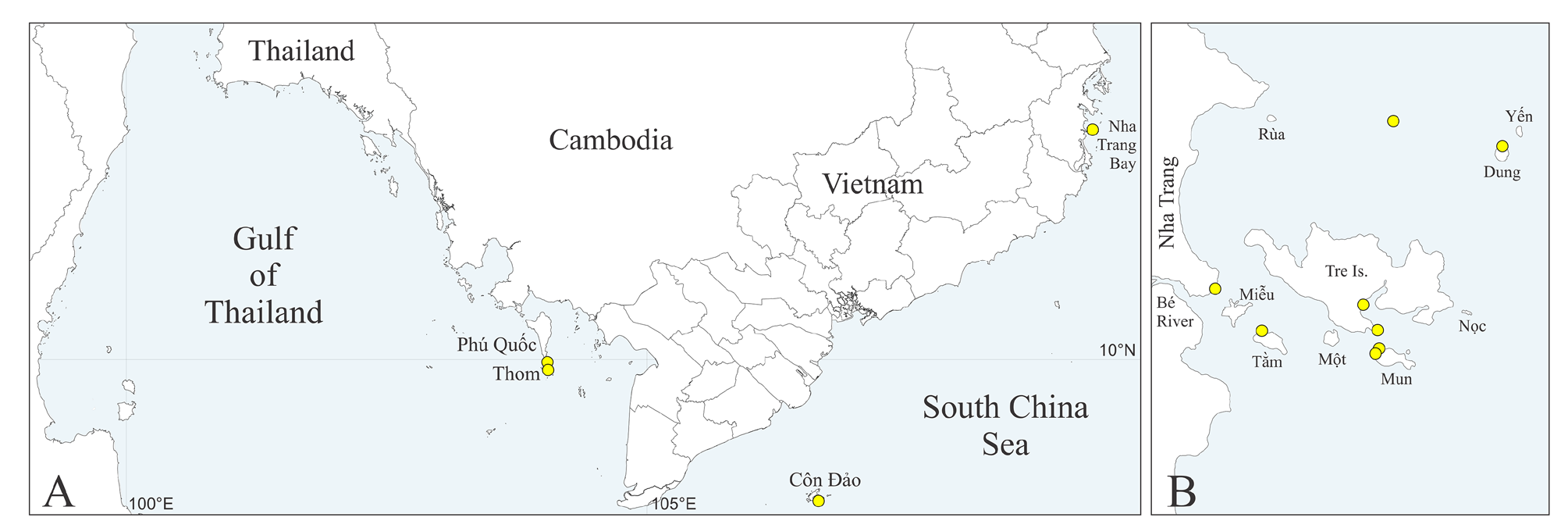
Type material. South-China Sea, Vietnam, Gulf of Thailand, An Thoi Archipelago, Thom Is., 5 m, in shell of the gastropod Vasum turbinellus (Linnaeus) occupied by hermit crab, 4 Feb 1986, MIMB 42722 View Materials (paratype) . Nha Trang Bay: Tam Is., 10 m, in dead coral encrusted by coralline alga, 22 May 2006, MIMB 42723 View Materials (4 paratypes); Dung Is., 10 m, in shell of Chama sp. , 2 Jun 2006, MIMB 42724 View Materials (26 paratypes); Tre Is., eastern side, 15 m, in dead coral, 14 Jun 2006, MIMB 42725 View Materials (40 paratypes), 42726 (holotype); Nha Trang, pier fouling, 3 m, in coral on pillar, 11 Jul 2006, MIMB 42727 View Materials (25 paratypes); under crust of coralline alga on shell of Arca avellana Lamarck bought in Nha Trang market, 30 Apr 2021, MIMB 42728 View Materials (5 paratypes); in oyster shell bought in Nha Trang market, 30 Apr 2021, MIMB 42729 View Materials (paratype) . Complete data on material examined is provided in Supplementary Table S3.
Adult morphology. Adults up to 21 mm long, 0.55 mm wide for 125 chaetigers. Pigmentation absent on body; some individuals with fine black pigment diffused on sides of palp grooves. Prostomium anteriorly bifurcated, posteriorly extending to end of chaetiger 4 as a low caruncle, shorter in small individuals ( Figs 11A View FIGURE 11 , 12A View FIGURE 12 ). Eyes and occipital antenna absent. Palps as long as 15−20 chaetigers, with frontal longitudinal groove lined with fine cilia, latero-frontal motile compound cilia on sides of groove, short compound non-motile cilia arising directly from palp surface and scattered on lateral and abfrontal palp surfaces.
Chaetiger 1 with short capillaries and small postchaetal lamellae in both rami. Posterior notopodia with only capillary chaetae.
Chaetiger 5 larger than chaetigers 4 and 6, with up to five dorsal superior capillaries ( Fig. 13F View FIGURE 13 ), five falcate spines alternating with bilimbate-tipped companion chaetae ( Fig. 13 View FIGURE 13 A−D) and arranged in oblique slightly curved row, and six ventral capillaries ( Fig. 13E View FIGURE 13 ). Dorsal superior and ventral capillaries winged, shorter and fewer than those chaetae on chaetigers 4 and 6. Falcate spines with spoon-like hollow on subdistal concave side ( Fig. 13B, D View FIGURE 13 ).
Hooded hooks in neuropodia from chaetiger 7, up to five in a series, accompanied by 1−2 winged inferior capillaries until chaetigers 9−10 ( Fig. 13G View FIGURE 13 ), and alternating with 1−2 hair-like alimbate capillaries in 5−10 posterior neuropodia ( Fig. 13I View FIGURE 13 ). Hooks bidentate, with slightly curved shaft without constriction.
Branchiae usually from chaetiger 9, occasionally from chaetigers 8 or 10, full-sized from chaetigers 12−14, very short after middle of body and absent from posterior 1/3−1/8 part of body ( Fig. 12B View FIGURE 12 ), free from notopodial postchaetal lamellae, flattened, with surfaces oriented parallel to body axis.
Nototrochs from chaetigers 6−7 in small individuals with up to about 40 chaetigers, from chaetigers 10−16 in larger worms, composed of single rows of ciliated cells in both sexes. Intersegmental ciliation absent.
Pygidium usually thin, flaring, disc-like to cup-shaped, up to two times wider than posterior most chaetigers, trilobed, with one ventral lobe and two smaller dorsal lobes, white due to numerous fusiform glandular cells. Lateral clefts shallow and indistinct in some individuals, thus pygidium resembling a cup or a disc with only dorsal incision ( Fig. 11D, E View FIGURE 11 ).
Glandular pouches in neuropodia from chaetiger 6, full-sized from chaetigers 7−8, diminishing in size after chaetigers 10−13.
Digestive tract with gizzard-like structure beginning from chaetigers 9−18 ( Fig. 12A View FIGURE 12 ) and extending through 1−2 chaetigers. Gizzard composed of anterior transparent muscular part and posterior white, apparently secretory, part. Rectum white in 3−5 posterior most chaetigers.
Nephridia from chaetiger 7, opening to exterior via two nephridiopores situated on lateral sides in anterior sterile chaetigers and on dorsal side in fertile chaetigers.
MG staining. Weak staining appeared on the lateral and ventral sides of the peristomium; the foregut was evenly colored ( Fig. 11F, G View FIGURE 11 ). Paired blotches (gatherings of strongly stained, probably glandular, cells) appeared on the dorsal side from chaetigers 19–22 onwards ( Fig. 11C, F View FIGURE 11 ), except 10−20 posterior most chaetigers. Narrow transverse bands were weakly stained on the ventral side of chaetigers 6 and 11; wider bands of stronger staining were present on chaetigers 8–10 ( Fig. 11B View FIGURE 11 ). In most specimens, peristomium and ventral side of chaetigers 6 and 11 were not stained, whereas in some specimens the dye was not absorbed on the ventral side of anterior chaetigers. In 20 of 21 stained specimens, the prostomium and about 20 anterior chaetigers did not absorb the dye ( Fig. 11F View FIGURE 11 ), whereas in one specimen intense staining appeared on the dorsal side of prostomium and eight anterior chaetigers ( Fig. 11A View FIGURE 11 ). Glandular pouches in neuropodia absorbed the dye and were well seen in lateral view from chaetigers 8−11 to chaetigers 25−30 ( Fig. 11G View FIGURE 11 ).
Habitat. Dipolydora vietnamita sp. nov. is an opportunistic borer, making burrows in corals, coralline algae, shells of the jewel box clam Chama sp. , hazelnut ark Arca avellana Lamarck , and vase snail Vasum turbinellus (Linnaeus) occupied by hermit crabs. This species is common in the coastal waters of Vietnam, co-occurring with other shell-boring polydorins in the same shells and corals. Sometimes the worms occur in dense aggregations up to ten individuals per square centimeter, forming colonies of hundreds of worms in one shell and thousands of worms in a coral. Two worms were found boring in a shell of the date mussel Lithophaga sp. which, in turn, perforated the shell of the hazelnut ark Arca avellana ( Fig. 14 View FIGURE 14 A−C).
Reproduction. Dipolydora vietnamita sp. nov. is dioecious. Paired gonads are attached to segmental blood vessels from chaetigers 21−24 through 2/3−3/4 of the body. Spermatogonia proliferate in testes; spermatogenesis occurs in the coelomic cavity. Spermatids are joined in tetrads ( Fig. 13K View FIGURE 13 ). Spermatozoa are introsperm about 1 µm in diameter, with a pointed acrosome 3 µm long, nucleus 5 µm, middlepiece 4 µm, head+middlepiece 12 µm long, and flagellum 54 µm long. Egg morphology and larval development of the species remain unknown.
Asexual reproduction. Many fragments of worms regenerating anterior and posterior chaetigers were found in shells from Nha Trang Bay in June 2006. Examination of those fragments has allowed for determining the course of the architomic asexual reproduction of D. vietnamita sp. nov. In the beginning, individuals divide into two or three stock fragments, each of which subsequently regenerates lost parts. The smallest stock fragment had six middle chaetigers. The anterior regeneration starts with a bud that grows and differentiates into head and body anlage. Palps begin to develop immediately after head differentiation. The body anlage elongates gradually and becomes segmented, probably simultaneously, into eight segments; no more segments are added anteriorly. Chaetae, postchaetal lamellae and other external segment attributes, as well as mouth and foregut, develop only when the segmentation is completed. Eyes never develop in the regenerated head. The posterior regeneration also starts with a bud that gives rise to the pygidium and the pre-pygidial growth zone. New posterior stolon segments develop successively following the formation of the growth zone. Similar records of architomy have been found in other spionids such as Pygospio elegans Claparède, 1863 ( Rasmussen 1953) and Dipolydora armata ( Radashevsky & Nogueira 2003) .
Remarks. Dipolydora vietnamita sp. nov. is similar to D. socialis ( Schmarda, 1861) from Chile, and D. protuberata from New South Wales, Australia, by having subdistal protuberance on the falcate spines of chaetiger 5. However, the two latter species are tube-dwellers in soft sediments while D. vietnamita sp. nov. is a shell- and coral-borer. Dipolydora vietnamita sp. nov. and D. socialis have trilobed pygidia but differ in the arrangement of branchiae and composition of the gizzard-like structures. Dipolydora socialis has branchiae usually beginning from chaetiger 8 and gizzard with four chitinous plates attached to the inner wall of muscular part, while D. vietnamita sp. nov. has branchiae usually beginning from chaetiger 9 and gizzard without chitinous plates. Dipolydora vietnamita sp. nov. differs from D. protuberata by having trilobed pygidium instead of disc-like pygidium with a dorsal notch.
Etymology. The species name refers to the type locality of the species in Vietnam.
Distribution. South-China Sea, Vietnam (see Table S3, Fig. 1A, B View FIGURE 1 ).




| MG |
Museum of Zoology |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |